Ячеистые блоки стеновые: Газобетонный блок 600*300*200, Д400, 500, 600
Блоки из ячеистого бетона: характеристики и особенности
Блоки стенового ячеистого бетона являются одним из самых распространенных материалов для строительства малоэтажных домов. Уже несколько десятилетий эти материалы используются в строительных работах. Но о ячеистом бетоне необходимо знать больше, так как стеновой ячеистый бетон сложнее, чем кажется. И в этой статье мы расскажем вам о характеристиках, видах, условиях хранения и так далее.
Это искусственный каменный материал, который отличается от аналогов пористостью. Пористость придает ячеистому бетону легкость и высокие показатели сохранения тепла в помещении. Для его изготовления используется специальный известковый, цементный или смешанный составы.
Поры в ячеистом бетоне подразделяются на газовые, капиллярные и гелевые. В первом случае поры образуются из-за выделения водорода в процессе реакции при смешивании требуемых компонентов.
Современные ячеистые газобетонные блоки – это еще один шаг вперед в мире строительства. Они соответствуют строительным нормам и правилам по теплопроводности и прочности. Также характеристика материала определяет большую скорость возведения, благодаря точной геометрии и простоте кладки.
Классификация ячеистого бетона
Блоки ячеистого бетона подразделяются на множество видов, в зависимости от определяющих факторов и характеристик.
- По назначению стеновой ячеистый бетон бывает конструкционным, теплоизоляционным и смешанным.
- Характеристика твердения бетона определяет автоклавные (блок твердеет под воздействием пара и повышенного давления) и неавтоклавные (блок твердеет в обычных условиях) материалы.
-
Поры образуются несколькими способами, что характеризует газобетоны, пенобетоны, аэрированные и смешанные виды.

- Для изготовления стенового ячеистого бетона используются следующие составляющие: известь, сланец, шлак и цемент.
- В качестве заполнителя выступают вторичные промышленные продукты и песок.

Как правильно хранить ячеистые блоки
Технические характеристики стеновых ячеистых бетонных блоков определяют условия хранения. Важно следовать им, чтобы материал сохранил все полезные свойства. На объект привозят материал, который необходимо разгружать ровно блок за блоком на специальных поддонах. Важно исключить перекосы и возможность попадания влаги. В сухую погоду каждый блок необходимо увлажнить, чтобы материал не пересох. Характеристика этого стенового материала предполагает внимательность и особый уход при хранении.
Стеновой ячеистый бетон: надежность под вопросом?
Стеновой ячеистый бетон известен своей хрупкостью. Эта характеристика не позволяет обращаться с ним так же, как с кирпичом.
Однако это может быть нивелировано с использованием монолитного пояса. Это особое устройство, обеспечивающее дополнительную жесткость конструкции и принимающее на себя вертикальные нагрузки. Монолитный пояс равномерно распределяет нагрузку между несущими стенами. Применение монолитного пояса в ряде случаев является необходимостью из-за особенностей этого стенового материала. Монтируют монолитный пояс, как правило, на уровне межэтажного перекрытия. Таким образом, стеновой ячеистый бетон не уступает альтернативным материалам в соотношении цена-качество.
Оформить заказ
Если вы хотите приобрести стеновой ячеистый бетон, то вам достаточно позвонить по указанному номеру телефона или заказать товар через сайт. Мы всегда готовы помочь вам с выбором и оформить заказ на стеновой ячеистый бетон в Москве и других городах России.
Мы всегда готовы помочь вам с выбором и оформить заказ на стеновой ячеистый бетон в Москве и других городах России.
Пенобетон производится из цемента, воды, песка и пенообразователя. Есть две технологии его производства. Литиевая, при которой исходная масса заливается в формы определенных размеров. И резательная, при которой изготавливается плита большого размера, которая потом разрезается на пеноблоки нужных габаритов.
Блоки стеновые мелкие из ячеистых бетонов. Согласно ГОСТу 21520-89
Блоки стеновые мелкие из ячеистых бетонов. Маркировка, транспортировка и хранение. Согласно ГОСТу 21520-89
Каковы основные параметры и размеры стеновых мелких блоков из ячеистых бетонов?Ответ: Стеновые мелкие блоки из ячеистых бетонов предназначены для кладки стен и перегородок жилых, общественных и производственных зданий с относительной влажностью помещений не более 75%.
Согласно ГОСТу 21520-89, типы стеновых мелких блоков должны соответствовать показателям, указанным в таблице.
Табл. Типы и размеры стеновых мелких блоков из ячеистого бетона
|
Типы стеновых мелких блоков |
Размеры для кладки (мм) |
|||||
|
|
на клею |
|||||
|
высота |
толщина |
длина |
высота |
толщина |
длина |
|
|
|
188 |
300 |
588 |
198 |
295 |
598 |
|
II |
250 |
245 | ||||
|
200 |
195 |
|||||
|
III |
288 |
298 |
||||
|
IV |
188 |
388 |
198 |
|
||
|
288 |
288 |
298 |
298 |
|||
|
V |
250 |
245 |
||||
|
VI |
|
300 |
588 |
— |
— |
— |
|
VII |
119 |
250 |
||||
|
VIII |
88 |
300 |
98 |
295 |
598 |
|
|
IX |
250 |
245 |
||||
|
X |
200 |
398 |
195 |
398 |
||
Ответ: Согласно ГОСТу 21520-89, марки бетона по морозостойкости должны быть в зависимости от режима их эксплуатации и расчетных зимних температур наружного воздуха в районах строительства, не менее:
— F25 — для блоков наружных стен;
— F15 — для блоков внутренних стен.
Ответ: Согласно ГОСТу 21520-89, классы (марки) бетона по прочности на сжатие и марки бетона по средней плотности должны быть не ниже класса (марки) по прочности
В1,5 (М25) и марки по средней плотности не выше D1200.
Какая должна быть отпускная влажность бетона мелких стеновых блоков из ячеистых бетонов?Ответ: Согласно ГОСТу 21520-89, Отпускная влажность бетона блоков не должна превышать (по массе) более, %:
-25 — на основе песка;
— 35 — на основе золы и других отходов производства.
Как расшифровывается условное обозначение мелких стеновых блоков из ячеистого бетона?Ответ: Условное обозначение мелких стеновых блоков из ячеистого бетона должно состоять из следующих обозначений:
— тип блока;
— класс (марка) бетона по прочности на сжатие;
— марка по средней плотности;
— марка по морозостойкости;
— категория.
Пример: I-В2,5D500F35-2 – блок типа I, класса по прочности на сжатие В2,5, марки по средней плотности D500, марки по морозостойкости F35 и категории 2.
Как маркируются мелкие стеновые блоки из ячеистого бетона отпускной партии товара?Ответ: Согласно ГОСТу 21520-89, товарные партии мелких стеновых блоков отличающиеся марками бетона по средней плотности и классами по прочности, следует маркировать несмываемой краской.
Маркировку следует наносить не менее чем на два блока (с противоположных сторон контейнера или пакета) цифрами, обозначающими среднюю плотность бетона блоков и класс по прочности на сжатие. Для блоков с маркой бетона по средней плотности от D500 до D900 следует наносить одну первую цифру числа, от D1000 до D1200 — две первые цифры числа, например: если блоки в партии имеют марку бетона по средней плотности D600 и класс по прочности на сжатие В2,5, то на блоки наносят цифры 6-2,5.
При марке бетона по средней плотности D1000 и классе по прочности на сжатие В7,5 наносят цифры10-7,5.
На каждое упакованное место должен быть нанесен знак «Боится влаги» по ГОСТ 14192.
Как соотносятся между собой марки бетона по средней плотности и класс бетона по прочности на сжатие для стеновых мелких блоков из ячеистого бетона?Ответ: Соотношение марок бетона по средней плотности с классами бетона по прочности на сжатие зафиксировано ГОСТом 21520-89 и приведено в табличную форму.
Табл. Соотношение марки бетона по средней плотности и класса бетона по прочности на сжатие
|
Марка бетона по средней плотности |
D500 |
D600 |
D700 |
D800 |
D900 |
D1000 |
D1100 |
D1200 |
|
Класс бетона по прочности на сжатие блоков из бетонов неавтоклавного твердения, не менее |
В3,5 |
В5 |
В5 |
В7,5 |
В7,5* |
В7,5* |
В10* |
В12,5* |
|
В2,5 |
В3,5 |
В3,5 |
В5 |
В5* |
В5* |
В7,5* |
В10* |
|
|
В2 |
В2,5 |
В2,5 |
В3,5 |
В3,5* |
— |
— |
— |
|
|
В1,5 |
В2 |
В2* |
В2,5 |
В2,5* |
— |
— |
— |
|
|
* Показатели класса по прочности на сжатие относятся только к блокам из бетона неавтоклавного твердения. |
||||||||

Ответ: Согласно ГОСТу 21520-89, мелкие стеновые блоки из ячеистого бетона должны перевозиться на поддонах с жесткой фиксацией термоусадочной пленкой, обеспечивающей неподвижность и сохранность изделий, а так же в специальных контейнерах.
Перевозка мелких стеновых блоков из ячеистого бетона разрешена любым видом транспорта при условии выполнения требований ГОСТ 9238 и Технических условий погрузки и крепления грузов.
Запрещено производить загрузку и транспортировку стеновых мелких блоков из ячеистого бетона навалом и разгрузку сбрасыванием.
Хранить стеновые мелкие блоки необходимо в штабелях высотой не более 2,5 метра.
виды, характеристики, технология укладки, цены
Строительные блоки из ячеистого бетона ценятся за высокие тепло- и звукоизоляционные свойства, малый вес и быстроту кладки. Уникальная пористая структура поддерживает нужную температуру внутри помещения, материал легко поддается обработке. Для изготовления используются особые технологии, благодаря которым блоки имеют правильную геометрию с четко заданными размерами, ровные стенки и достаточно высокую прочность. Это доступный материал, его можно купить в любом строительном супермаркете или заказать в интернет-магазине.
Уникальная пористая структура поддерживает нужную температуру внутри помещения, материал легко поддается обработке. Для изготовления используются особые технологии, благодаря которым блоки имеют правильную геометрию с четко заданными размерами, ровные стенки и достаточно высокую прочность. Это доступный материал, его можно купить в любом строительном супермаркете или заказать в интернет-магазине.
Оглавление:
- Отличия от других видов
- Разновидности и их технические параметры
- Где используются?
- Достоинства и недостатки
- Особенности монтажа
- Цены
Особенности и свойства
Данная разновидность легкого бетона имеет минеральную основу, наполнителем служат кремнеземистые порообразующие порошки. В качестве вяжущих компонентов выступают цемент и известь (редко — гипс), также в состав вводятся: зола, отходы металлургического производства, кварцевый песок. Для образования закрытых ячеек применяют различные технологии, благодаря которым общая пористость материала достигает 85–90 % при надежной базе.
Закрытая структура защищает ячеистый бетон от влаги и агрессивных внешних воздействий, а минеральная основа делает его стойким к грибку и плесени. Такой стройматериал имеет малую плотность и вес и характеризуется уникальными тепло- и звукоизоляционными свойствами.
Виды, размеры и характеристики
Классификация и нюансы эксплуатации определяются по ГОСТ 25485-89 «Бетоны ячеистые. Технические условия». Основное различие касается способа поризации, выделяют газобетон, аэрированный ячеистый силикат и пенобетон, последний получают путем введения в смесь специальной пены. Изготовленные с помощью газообразования стеновые блоки имеют самую высокую прочность, их физико-механические свойства одинаковы по всему объему. Структура пенобетона более рыхлая, отклонения в размерах достигают 5 мм, ячейки, расположенные на поверхности недостаточно закрыты и подвержены внешним воздействиям. Газосиликатные блоки характеризуются высокой точностью форм, погрешность в линейных размерах лежит в пределах 1–2 мм. В зависимости от способа застывания различают: пропариваемые, прогреваемые и изделия автоклавного твердения, последние ценятся за гладкую и защищенную поверхность.
Газосиликатные блоки характеризуются высокой точностью форм, погрешность в линейных размерах лежит в пределах 1–2 мм. В зависимости от способа застывания различают: пропариваемые, прогреваемые и изделия автоклавного твердения, последние ценятся за гладкую и защищенную поверхность.
Функциональное назначение определяется плотностью объемной массы: для теплоизоляции хватает 300–500 кг/м³, для возведения строительных конструкций — от 500 до 900 (при условии обязательного армирования), для многоэтажных жилых, промышленных и сельскохозяйственных объектов — 1000–1200 и более. Последняя разновидность может использоваться в качестве перекрытия, размер обычно стандартный (600×200×250 мм). Существует прямо пропорциональная связь между плотностью и прочностью на сжатие, теплопроводностью и морозостойкостью (чем она выше, тем качественнее бетон).
По целевому назначению различают стеновые ячеистые изделия, перегородки, перекрытия и перемычки. Их габариты обычно стандартные, эксплуатационные характеристики также определяются ГОСТ 25485-89. В основном они отличаются по длине, минимальные размеры стеновых мелких блоков составляют 400×200×200 мм, перегородочных 300×400×120. В свою очередь максимальные: 600×500×250 мм и 600×400×150, соответственно. Предлагаемый ассортимент весьма широк, в продаже есть элементы с нестандартной длиной (625 мм) или шириной (375, 288).
В основном они отличаются по длине, минимальные размеры стеновых мелких блоков составляют 400×200×200 мм, перегородочных 300×400×120. В свою очередь максимальные: 600×500×250 мм и 600×400×150, соответственно. Предлагаемый ассортимент весьма широк, в продаже есть элементы с нестандартной длиной (625 мм) или шириной (375, 288).
Сфера применения
Согласно ГОСТ 25485-89, стеновые блоки из ячеистого бетона рекомендуется выбрать для возведения наружных утепляющих панелей жилых домов и общественных комплексов или внутренних несущих конструкций и перегородок. Также они пригодны для теплоизоляции чердачных перекрытий, промышленного оборудования с температурой поверхности до 700 °C, трубопроводов — до 400.
Простота обработки и правильная геометрия блоков делают их незаменимыми при необходимости быстрого возведения загородных многоэтажных коттеджей, возможна реализация сложных архитектурных решений. Допускается ведение строительства на грунтах с малой несущей способностью, нагрузка на фундамент снижается, по сравнению с другими видами стройматериалов. Альтернативой варианту использования ячеистого бетона является изготовление плит и сэндвич-панелей.
Альтернативой варианту использования ячеистого бетона является изготовление плит и сэндвич-панелей.
Плюсы и минусы
К преимуществам относят:
1. Огнестойкость и пожаробезопасность: стена толщиной в 10 см выдерживает прямое воздействие пламени в течение 2 часов, без потери своих полезных характеристик.
2. Низкую теплопроводность, по утепляющим свойствам ячеистая структура не уступает древесине или кирпичу и даже превосходит их.
3. Экономичность: стеновые блоки не нуждаются в дополнительной теплоизоляции при условии использования специальных клеевых составов при укладке. Расход смеси при этом минимальный, благодаря правильной геометрии и гладкой поверхности, отклонения не превышают 2–3 мм, как следствие — нет необходимости в толстом кладочном шве.
4. Малый вес крупногабаритных блоков.
5. Обрабатываемость: ячеистый бетон легко распилить на части или просверлить.
6. Устойчивость к биологическим воздействиям.
7. Отсутствие мостиков холода между блоками, при условии использования специальных вяжущих строительных смесей.
8. Высокую паропроницаемость, поддержание влажности в помещении на комфортном уровне.
9. Доступную стоимость блока.
10. Долговечность кладки (до 75 лет).
Недостатком является относительная хрупкость (малая прочность на излом), и, как следствие — необходимость в надежном основании. Для пеноблока огромное значение имеет подвижность грунта, рекомендуемый тип фундамента — столбовой или монолитный ленточный. При малом весе блоков обязательна организация цоколя из более тяжелого и плотного бетона, что сказывается на стоимости проекта, исключение касается лишь одноэтажных построек на устойчивых почвах. Требования к фундаменту при выборе газосиликатного бетона менее строгие.
Также материал нуждается в определенной защите от воздействий окружающей среды, обязательно грунтование и обработка специальными химреагентами. Даже ячеистый бетон автоклавного твердения разрушается в процессе эксплуатации без проведения облицовки (несмотря на заявления производителей об обратном). Хорошие отзывы о качестве защиты имеет фасадная краска или легкие наружные штукатурные смеси. Материал рассчитывается и закупается заранее, соблюдаются вентиляционные и влажностные нормативы: при длительном хранении полиэтиленовая пленка прорезается по бокам, блоки из бетона располагают на поддонах и увлажняются при пересушивании.
Хорошие отзывы о качестве защиты имеет фасадная краска или легкие наружные штукатурные смеси. Материал рассчитывается и закупается заранее, соблюдаются вентиляционные и влажностные нормативы: при длительном хранении полиэтиленовая пленка прорезается по бокам, блоки из бетона располагают на поддонах и увлажняются при пересушивании.
Нюансы кладки
Строительство начинается с размещения первого ряда на фундамент, рекомендуется вести кладку с угла, рядами по всему периметру будущего здания. Строго отслеживается уровень: допустимое отклонение составляет 30 мм, для контроля натягивается шнур, применяются отвесы и лазерные координаторы. Проверяется жесткость фундамента, обеспечивается максимально качественная гидроизоляция (при попадании влаги внутрь и последующем промерзании грунта возрастает риск смещения кладки и растрескивания стеновых блоков). Рулонные материалы размещаются с обязательным нахлестом, основание обрабатывается гидрофобными битумными мастиками.
Перед началом кладки блоки проверяются на целостность, поврежденные используются для распила. Размеры швов имеют значение: при выборе цементного раствора их толщина варьируется по горизонтали от 10 до 15 мм, по вертикали — от 8 до 15, оптимальными считаются средние показатели. Во избежание образования мостиков холода все возможные пустоты заполняются, более экономным вариантом является монтаж на клей, толщина швов составляет 2±1 мм. Для обеспечения надежности постройки проводится укладка металлических стержней на раствор между стеновыми блоками. Обязательно армируется нижний ряд, каждый четвертый, зоны оконных проемов и перемычек, участки с высокой нагрузкой. Для увеличения полезной площади дома допускается смещение возводимой конструкции, но не более чем на треть от общей величины.
Размеры швов имеют значение: при выборе цементного раствора их толщина варьируется по горизонтали от 10 до 15 мм, по вертикали — от 8 до 15, оптимальными считаются средние показатели. Во избежание образования мостиков холода все возможные пустоты заполняются, более экономным вариантом является монтаж на клей, толщина швов составляет 2±1 мм. Для обеспечения надежности постройки проводится укладка металлических стержней на раствор между стеновыми блоками. Обязательно армируется нижний ряд, каждый четвертый, зоны оконных проемов и перемычек, участки с высокой нагрузкой. Для увеличения полезной площади дома допускается смещение возводимой конструкции, но не более чем на треть от общей величины.
Стоимость
| Наименование марки ячеистого бетона, производитель | Назначение блоков | Класс прочности | Плотность, кг/м3 | Тепло- проводность, Вт/м∙С | Размеры, мм | Цена за 1 м3, рубли |
| Poritep, Рязань | стеновые | В3,5 | 500 | 0,11 | 625×250×200 | 3 500 |
| 625×250×175 | 3 600 | |||||
| Ytong D500, Xella, Германия
| 0,099 | 625×500×250 | 4 770 | |||
| U-образные, перемычка | 500×300×250 | 380 за 1 шт | ||||
| Драубер, Электросталь | перегородка | В2,5 | 0,12 | 600×200×400 | 3 670 | |
| ДСК Грас, Калуга | стеновые | В3,5 | 600 | 0,14 | 625×300×250 | 3 800 |
| перегородка | 500 | 0,12 | 625×200×250 | 3 720 | ||
| КЗСМ, Кострома | стеновые | В2,5 | 400 | 0,11 | 600×500×250 | 3 550 |
| В3,5 | 500 | 0,13 | 600×450×250 | 3 600 |
Стеновые блоки из ячеистых бетонов
ГОСТ 21520-89 «Блоки из ячеистых бетонов стеновые мелкие»
Главная|ГОСТы и СНиП|ГОСТ 21520-89 «Блоки из ячеистых бетонов стеновые мелкие»
Дата: 30 декабря 2016
Просмотров: 867
Коментариев: 0
Строительные технологии совершенствуются день ото дня. Касается это, в первую очередь, материалов, применяемых при возведении зданий различного назначения. Искусственные композиты не спеша отодвигают на второй план, привычные всем, кирпич и железобетонные панели. Одну из лидирующих позиций в списке занимают блоки из ячеистого бетона. Что же представляет собой этот материал, производство которого осуществляется в соответствии с требованиями стандарта?
Касается это, в первую очередь, материалов, применяемых при возведении зданий различного назначения. Искусственные композиты не спеша отодвигают на второй план, привычные всем, кирпич и железобетонные панели. Одну из лидирующих позиций в списке занимают блоки из ячеистого бетона. Что же представляет собой этот материал, производство которого осуществляется в соответствии с требованиями стандарта?
Общие понятия
Ячеистый бетон относится к группе легких бетонов. Благодаря большому количеству искусственно созданных пор (примерно 85%), он при относительно небольшой массе имеет отличные теплоизоляционные и прочностные характеристики. Одним из положительных факторов, определяющих приоритетное положение ячеистых материалов в общем списке, является правильная геометрическая форма. Отклонения размеров не превышают 2мм, что значительно облегчает ход строительных мероприятий. Возведение стен производится при помощи специального клея, не создающего, так называемых, мостиков холода.
Блоки из ячеистого бетона – отличная альтернатива кирпичной кладки
К положительным свойствам ячеистых композитов можно отнести:
- Высокие прочностные характеристики, позволяющие использовать блоки для возведения несущих стен в зданиях высотой до 15 метров.
- Возможность выполнения теплоизоляции оборудования, поверхность которого нагревается до 700 ⁰С.
- Отсутствие возможности образования плесени и гнили, размножения грибков.
- Морозоустойчивость, позволяющая использовать материал в климатических зонах с низкими температурами.
- Влагоустойчивость, которая достигается благодаря замкнутой структуре пор.
- Высокая экологичность, которая снимает вопросы об отрицательном воздействии стройматериалов на человека.
- Долговечность, благодаря которой можно не беспокоиться о надежности возводимого объекта.
- Пожарную безопасность, так как материал не подвержен горению.
- Высокую податливость, позволяющую легко обрабатывать изделия и придавать им не стандартную конфигурацию.
- Снижение усилий на основание здания за счёт применения изделий, обладающих небольшой массой, несмотря на значительные габариты.
- Отсутствие необходимости в специальном грузоподъемном оборудовании для подъема композита к месту работ, что значительно уменьшает сроки выполнения строительных мероприятий.
- Повышенный уровень звуковой изоляции, связанный с высоким поглощением звуков, благодаря ячеистой структуре массива.


Разновидности ячеистого бетона
Для обеспечения всех положительных свойств производство блоков должно проводиться в строгом соответствии ГОСТ под номером 21520, разработанным и изданным в 89-м году.
Данный материал обладает целым рядом преимуществ, благодаря которым широко применяется для строительства жилых домов
В строительстве используются блоки из ячеистого бетона трех основных видов:
- пенобетон, получаемый путем смешивания специально приготовленной пены с цементным раствором;
- газобетон, образование пор в котором происходит в результате химической реакции реагента с основным составом;
- газосиликат, поры в котором получаются за счет реакции извести, измельченного песка, воды с газообразующими компонентами.
На данные виды бетона разработана техническая документация, требующая соблюдения технологического процесса при производстве. Она регламентирует допускаемые отклонения геометрических параметров, правила монтажа готовых изделий.
Она регламентирует допускаемые отклонения геометрических параметров, правила монтажа готовых изделий.
Сфера применения
Широкая сфера применения ячеистых композитов обусловлена высокими теплоизоляционными характеристиками. Это позволяет задействовать их при выполнении таких задач, как:
- Утепление стеновых и потолочных железобетонных перекрытий.
- Теплоизоляция чердачных помещений, что является особо актуальным при возведении зданий с мансардой, предполагающих возможность круглогодичного проживания в них.
- Создание термопрокладки при строительстве многослойных конструкций. В данном случае обеспечивается комфортный температурный режим помещений, без резких колебаний.
- Обеспечение теплоизоляции оборудования, имеющего повышенные температурные показатели поверхности.
- Создание теплового барьера при укладке трубопроводов.
- Возведение стен частных домов и объектов небольшой этажности.
Для того чтобы получить ячеистый бетон механическим способом, делается раствор на основе цемента, в который добавляется песок и вода
Рассмотрим подробнее, какие требования на блоки из ячеистых бетонов стеновые оговорены в действующих нормативных документах.
Основные контролируемые параметры
Действующий ГОСТ на стеновые мелкие композиты оговаривает следующие моменты:
- сферу применения;
- технические особенности;
- специфику приемочного контроля;
- методику проверки;
- условия хранения и транспортировки.
Область использования
Блоки из ячеистых бетонов стеновые применяются при возведении внешних стен, а также внутренних перегородок. Не рекомендуется возведение объектов в местах с повышенной влажностью, превышающей 75%. Использование ячеистых материалов при влажности воздуха выше 60% возможно, при условии, что их внутренняя поверхность будет покрыта пароизолирующим слоем.
Очень выгодно и экономически оправдано сооружение из ячеистых материалов построек частного типа: коттеджей, офисов, садовых домиков, складских помещений, гаражей. При возведении несущих стен допускается применение, если высота здания не превышает 15-ти метров, при самонесущих стенах – не превышающих 30-ти метровой высоты.
Использование стеновых блоков из ячеистого бетона при строительстве обеспечивает высокую пожаробезопасность, герметичность стен, полов, перегородок
Технические параметры
В требованиях нормативного документа четко оговорены габариты и различные особенности. Стандарт осуществляется разбивку стеновых изделий из ячеистых композитов на десять типов.
Согласно типам композита, а также возможности кладки с помощью раствора, их геометрические размеры изменяются следующим образом:
- Длина композитов стандартизирована и представляет типоразмерный ряд от 288 до 588 мм.
- Ширина представлена линейкой размеров в диапазоне 88-288 мм.
- Толщина составляет 200, 250, 300 мм.
Документ предусматривает возможность фиксации изделий из ячеистого бетона с помощью клея. При этом геометрические размеры для каждого типа отличаются от указанных выше и представлены следующим типоразмерным рядом:
- длина составляет 298, 398, 598 мм;
- ширина изменяется следующим образом: 195, 245,295 мм;
- высота композитов составляет 98, 198, 298 мм.

Разрешается в соответствии с заявками потребителей производить изготовление блоков, отличающихся размерами. Для продукции одинаковых типов ее толщина при фиксации на клей меньше, чем допускаемое значение ширины при установке с помощью раствора.
Настоящий стандарт распространяется на стеновые мелкие блоки из ячеистых бетонов
В обязательном приложении к стандарту уточняется, какая марка бетона может использоваться для производства:
- Составы, маркируемые D500, D600 и D700 применяются композитов с 1 по 10-й.
- Смесь D800 соответствует 2, 3, а также 5-10 видам.
- Раствор D900 приемлем для 3 и 5-10 типов.
- Бетон плотностью D1000 можно использовать для изделий 5-10 видов.
- Состав D1100 соответствует 5,6, а также 8-10 классам.
- Бетон со средней плотностью D1200 применим для 5 и 10 типов.
При использовании различных марок бетона не рекомендуется нарушать предусмотренные нормативным документом соотношения.
Особенности маркировки
Производя изготовление продукции, ее шифр при заказе должен включать следующую информацию:
- маркировку типа изделия;
- марку или классификацию бетонного состава, характеризующую его прочность;
- цифровой индекс, обозначающий удельный вес бетонного массива;
- обозначение устойчивости массива к отрицательным температурам;
- категорию.

Материалы и бетон для изготовления блоков должны соответствовать требованиям
Рассмотрим конкретную маркировку блока II-В7,5D800F35-3 и расшифруем ее параметры:
- II – характеризует типоразмер.
- В7,5 – расшифровывает класс изделия по его прочности и способности воспринимать сжимающие нагрузки.
- D800 – соответствует значению средней плотности бетонного состава.
- F35 – обозначает степень морозостойкости.
- 3 – индекс, характеризующий обозначение категории.
Любая партия товара, которая имеет отличие по средней плотности бетонного состава и классу прочности, в обязательном порядке должна маркироваться краской, которую невозможно смыть.
При расположении продукции в специальном контейнере или пакете маркировка должна выполняться с двух противоположных сторон упаковки. Например, если нанесено обозначение 8-7,5, то оно обозначает, что изделия в данной партии произведены из бетона, имеющего среднюю плотность D800, характеризуются классом прочности на сжатие В7,5.
Наличие данной информации позволяет заказчикам четко определить вид приобретаемой продукции, а производителю осуществить ее хранение и отгрузку в соответствии с указанным на упаковке типоразмером.
Общие характеристики
Прочностные характеристики применяемых бетонов не должны быть ниже В1,5, что соответствует марке М25, а также составу, удельный вес которого обозначается D1200.
Ячеистые бетонные блоки считаются одним из самых лучших строительных материалов, с их помощью можно возводить как жилые, так и хозяйственные помещения
Документ регламентирует критерии устойчивости блоков к воздействию отрицательных температур, что соответствует следующей маркировке:
- F25, если продукция применяется для внешних стен здания.
- F15 при использовании композитов внутри помещения.
Зависимость маркировки бетонного состава по среднему удельному весу и классификация растворов, согласно их твердости, приведены в таблице нормативного документа следующим образом:
- значению D500 соответствуют следующие классы бетона В1,5-В3,5;
- D600 и D700 – В2-В5;
- D800 и D900 – В2,5-В7,5;
- D1000 – В5, В7,5;
- D1100 – В7,5, В10;
- D1200 – В10, В12,5.

Показатели допусков
Качество блоков из ячеистого композита связано с отклонениями их геометрических параметров. Существует 3 различные категории ячеистых блоков, для которых документом предусмотрены значения предельных отклонений размеров:
- Первая категория, предназначенная для установки с помощью клея, отличается отклонением длины и толщины изделия ±2 миллиметра, а также высоты ±1 миллиметр. При этом допуск разности длины диагонали, характеризующий соответствие блока прямоугольной конфигурации составляет максимум 2 миллиметра.
Газобетонные блоки марок D600 и D500 с успехом используют в качестве теплоизоляционного и конструкционного материала
- Изделия 2-й категории устанавливаются на раствор и отличаются увеличенным допуском по длине и толщине, равным ±4 миллиметра, а также высоте ±3 миллиметра. Допускаемая разность диагоналей для них составляет 4 миллиметра.
- Товар 3-й группы, предназначенный для фиксации с помощью раствора, характеризуется максимальным значением допусков. Они составляют для высоты ± 5 миллиметров, а для длины и толщины блока ±6 миллиметров. Отклонение от прямоугольности имеет максимальное значение, равное 6 миллиметрам.
 Они составляют для высоты ± 5 миллиметров, а для длины и толщины блока ±6 миллиметров. Отклонение от прямоугольности имеет максимальное значение, равное 6 миллиметрам.
Они составляют для высоты ± 5 миллиметров, а для длины и толщины блока ±6 миллиметров. Отклонение от прямоугольности имеет максимальное значение, равное 6 миллиметрам.Помимо геометрических размеров изделия и отклонения от прямоугольной конфигурации, нормативный документ указывает возможную глубину повреждения ребер и углов блока. Она составляет от 5 до 15 мм в зависимости от категории продукции.
Схема измерения глубины повреждения стеновых блоков
Наибольшей степенью точностью характеризуются изделия первой категории, имеющие минимальное отклонение размеров.
Приемка продукции и контроль качества
Стандарт обязывает производителей продукции осуществлять приемку изделий, основываясь на результатах приемочного контроля и периодических испытаний. В процессе приемосдаточного контроля проверяются следующие параметры:
- габаритные размеры;
- способность продукции воспринимать сжимающие нагрузки;
- значение удельного веса;
- изменение влажности.

Документ регламентирует объем выборки для осуществления контроля, при обнаружении брака. Объем партии контролируемой продукции при повторных испытаниях удваивают. Любая партия товара сопровождается документом, подтверждающим его качество, с указанием в нем данных об организации, которая изготовила указанную продукцию.
Специфика складского хранения и доставки
Качественные показатели и сохранность продукции связаны не только с особенностями технологического процесса изготовления. Немаловажны особенности хранения и транспортировки. Документ предписывает осуществлять доставку продукции на специальных поддонах или контейнерах, обеспечивать ее неподвижность.
Запрещается отгружать россыпью и сбрасывать путем опрокидывания кузова. Хранение должно осуществляться с разбивкой по типоразмерам и категориям.
Заключение
Таковы основные положения стандарта на стеновые мелкие композиты, применяемые для изготовления блочных конструкций при возведении стен. Соблюдение требований нормативного документа гарантирует выпуск качественной продукции, соответствующий современным требованиям.
Соблюдение требований нормативного документа гарантирует выпуск качественной продукции, соответствующий современным требованиям.
pobetony.ru
Блоки из ячеистого бетона стеновые: характеристики и варианты использования
Материалы, предназначенные для возведения стен, нередко становятся объектом дискуссий среди застройщиков. Ведь выбор настолько велик, что принять решение становится непростой задачей.
Каждые изделия по-своему хороши, но и не лишены недостатков. Поэтому, необходимо предварительно изучить все положительные и отрицательные стороны материалов, и только потом принимать решение.
В данной статье мы поговорим о группе легких бетонов, а, точнее, ее представителях, которые в последнее время приобрели широкую популярность. Давайте разбираться, блоки из ячеистого бетона стеновые: чем они примечательны и каковы их основные характеристики?
Общее описание материала, особенности состава и технологии производства
Как уже говорилось, бетон ячеистый– это группа материалов. Представителями ее являются пено- и газобетон, а также пеногазобетон, сочетающий в себе обе технологии изготовления и, как следствие, свойства.
Представителями ее являются пено- и газобетон, а также пеногазобетон, сочетающий в себе обе технологии изготовления и, как следствие, свойства.
Теперь немного о составе. Он регулируется ГОСТ 21520-89 Блоки из бетона ячеистого стеновые мелкие.
- Такие блоки получают из смеси цемента, извести, песка, порообразователя, пластификатора и иных добавок, повышающих качества будущего материала.
- По сути, это смесь кремнеземистого компонента, жидкости, вяжущего и добавок, вызывающих вспучивание раствора и, как следствие, образование ячеек.
- В качестве кремнеземистого компонента могут выступать не только песок, но и зола высокоосновная, и иные отходы промышленности.
- Основным вяжущим, помимо цемента, может быть: шлак, смешанное вяжущее, зола, известь.
- Цемент, как правило, должен использоваться марки не ниже 400-500.
- Песок преимущественно добавляют кварцевый.
- Пластификаторы применяются с целью повышения пластичности раствора. В этом случае, блоки получаются лучшего качества. Они защищены от растрескивания на этапе производства.
- Если говорить о пеноблоке, то образование ячеек в растворе происходит за счет добавления пенообразователя. В случае с газоблоком, все несколько иначе. Поры образуются в следствие реакции алюминиевой пудры и извести негашеной.
 В этом случае, блоки получаются лучшего качества. Они защищены от растрескивания на этапе производства.
В этом случае, блоки получаются лучшего качества. Они защищены от растрескивания на этапе производства.- В результате структура пор у материалов разная: у пеноблока – закрытая, а у газоблока – открытая. Это также повлияло на некоторые значения свойств.
Обратите внимание! Соотношении пропорций при приготовлении смеси, напрямую влияет на итог. Например, более пористыегазо- или пеноблоки с небольшим содержанием цемента, будут менее тяжеловесными. Коэффициент теплопроводности у них будет низким, как и показатели прочности и плотности.
Повысить прочность можно при помощи некоторых приемов — вот несколько из них:
- Применение специализированных добавок;
- Использование в качестве кремнеземистого компонента песка, а не золы.
- Добавление большего количества вяжущего в процентном соотношении;
- Автоклавная обработка делает материал более прочным и устойчивым к механическим воздействиям.

Что касается процесса производства, то выпуск обоих видов блоков аналогичен.
- Первым делом замешивают раствор, который потом отправляется в формы.
- Наполняются они примерно на одну треть, так как вспучивание смеси может привести к перетеканию.
- После завершения процесса порообразования, продукция должна немного застыть.
- Далее производят распалубку.
- Последний этап зависит от того, какой метод твердения будет использован. Это может быть автоклавная обработка или сушка в естественных условиях.
Пено- и газобетон гидратационного твердения может быть произведен своими руками. Это значительно снизит цену строительства. Вышеуказанная инструкция вполне подойдет для применения.
Несколько слов об используемом оборудовании. В случае, если пено-, газоблоки производятся в заводских условиях, варианта может быть два: набор машин конвейерного типа либо стационарная линия оборудования.
В случае, если пено-, газоблоки производятся в заводских условиях, варианта может быть два: набор машин конвейерного типа либо стационарная линия оборудования.
В первом случае, производство будет максимально рентабельным, автоматизированным, объем продукции может достигать и 200, и 300 кубов в сутки.
Элемент линии конвейерного типаВо втором случае объем будет значительно меньше, да и вмешательство сторонних рабочих понадобится, а вот цена – более бюджетная.
Если блок стеновой изготавливается в домашних условиях, обойтись можно вполне комплектом из форм и смесителя. Также можно приобрести мобильную установку, с ней удастся произвести выпуск до 20 м3 в сутки.
Характеристика пено- и газоблока
С составом сырья и технологическим процессом мы немного ознакомились, теперь пришло время обратить внимание на основной набор характеристик материала и их числовые значения.
ГОСТ устанавливает ряд требований к материалу, которые обязаны соблюдать все производители. Однако в силу высокой конкурентной борьбы, каждый из них пытается выделить свою продукцию путем модернизации технологии изготовления и изменений пропорций сырья для повышения основных качеств. Давайте рассмотрим допустимый стандартом минимум, и предлагаемый изготовителями максимум.
Однако в силу высокой конкурентной борьбы, каждый из них пытается выделить свою продукцию путем модернизации технологии изготовления и изменений пропорций сырья для повышения основных качеств. Давайте рассмотрим допустимый стандартом минимум, и предлагаемый изготовителями максимум.
Технические и эксплуатационные свойства
Рассмотрим, используя таблицу, основные свойства двух представителей рассматриваемой группы материалов.
Таблица 1. Стеновые мелкие блоки из бетона ячеистого: технические характеристики:
| Наименование свойства | Значение его у пено-и газобетонного блока, комментарии |
| Морозостойкость | Показатель установлен ГОСТ. Минимальный порог – 25 циклов. Изделия, предназначенные для возведения наружных стен должны соответствовать данному значению в отношении марки морозостойкости. Для перегородочных блоков данное значение не установлено, также не нормируется морозостойкость теплоизоляционного вида материала. На практике же, производители уверяют потенциальных потребителей, что материал способен выдерживать до 150 циклов, а пеноблок – до 100. Подтверждения со стороны застройщиков пока еще не было, в силу недостаточного срока существования и применения блоков. |
| Средняя плотность, кг/м3 | Значение колеблется в промежутке от 300 до 1200. Как уже говорилось выше, плотность материала во многом определяет Серу его применения. В связи с этим, была разработана классификация материала в соответствии с показателем средней плотности. О ней мы поговорим чуть позже. |
| Прочность, марка В | Марки прочности должны соответствовать определенной плотности. Существующие находятся в промежутке от 1,5 до 15. |
| Теплопроводность Вт*мС | Теплоизоляционная способность достаточно высока. Коэффициент может равняться 0,08 и доходить до 0,34. При увлажненности во время эксплуатации, данное значение возрастет, однако это коренным образом не повлияет на теплоэффективность стены. |
| Экологичность | Состав мы уже рассмотрели и смогли убедиться в том, что смесь не содержит вредных и токсичных компонентов. |
| Пожароустойчивость | ГОСТ указывает на негорючесть материала. Более того, он может противостоять воздействию высокой температуры вплоть до нескольких часов. |
| Паропроницание | Оба представителя способны к паропроницанию. В помещении будет создаваться максимально благоприятный климат, так как блоки склонны впитывать лишнюю влагу и отдавать ее при преобладании сухого воздуха. |
| Звукоизоляция | Максимальной звукоизоляционной способностью обладают наименее прочные изделия. В целом, конструкционно-теплоизоляционного материала вполне достаточно для того, чтобы дополнительная звукоизоляция помещения не потребовалась. |

Разновидности блоков и их применение
Ознакомимся кратко с классификациями.
Выше мы уже упоминали о существовании нескольких способов твердения и говорили о том, что блок может твердеть посредством обработки его в автоклаве, либо естественным путем. В первом случае, воздействие оказывается при помощи давления, превышающего атмосферное и высокой температуры.
Во втором случае, возможна легкая тепловлажностная обработка, которая помогает повысить свойства и сократить процесс сушки.
Автоклав опережает своего конкурента во многих показателях, таких как:
- Марка морозостойкости и долговечность;
- Соотношение прочности, средней плотности и способности к теплосохранению;
- Блок автоклавного твердения имеет лучшую геометрию и чисто белый цвет;
- Он менее хрупкий.
В противовес стоит упомянуть о том, что стоимость продукции гидратационного твердения несколько ниже, примерно на 10%.
Материал синтезного и гидратационного тверденияПлотность положила начало появлению еще одной классификации.
- Блоки, характеризующиеся значением плотности в 300 или 400 кг/м2 называют теплоизоляционными. Применяют их, разумеется, в процессе теплоизоляции.
 Применяют их, разумеется, в процессе теплоизоляции.
Применяют их, разумеется, в процессе теплоизоляции.На заметку: Для этого используются не только блоки и плиты, но и монолитный материал. Он активно применяется при звуко- и теплоизоляции кровель, ограждающих конструкций, стяжке пола.
- Если изделия обладают значением плотности в 500, 600, 700, 800 или 900 (кг/м3), то они являются конструкционно-теплоизоляционными. С их помощью можно произвести возведение стены здания или перегородки. Они отлично подходят в качестве основного материала при малоэтажном строительстве.
- А вот если решено возводить 3- или 4-х этажную постройку, лучше приобрести газо- пеноблок с повышенным показателем прочности и плотности, то есть конструкционные. Они способны выдержать более значительную нагрузку, и с их помощью можно построить здание высотой в несколько этажей.
На заметку! В ассортименте пенобетона имеются также изделия, получившие название конструкционно-поризованные. Они – особенно плотные. Выпуск их в массовом количестве не производится, только лишь под заказ. ГОСТ об их существовании не упоминает.
ГОСТ об их существовании не упоминает.
Что касается назначения:
- Стены возводят при помощи стеновых блоков;
- Перегородки – с применением перегородочных;
- Стационарную опалубку можно соорудить из у-образных. Также их применяют при монтаже дверных и оконных проемов.
Блоки могут иметь различную категорию точности, которая зависит от величины допустимых отклонений. На фото ниже представлены требования стандарта.
Геометрические отклоненияОбратите внимание! От категории во многом зависит теплопроводимость будущей стены. Объясняется это очень просто. Например, продукцию первой категории укладывают на клей, толщина шва при этом соблюдается минимальная. Результат – минимум мостиков холода.
Если укладывать продукцию третьей категории, то применить клеевой состав не получится, необходимо использовать раствор, толщина слоя которого будет зависеть от отклонений от линейного размера. Достигать она может и 2-х см, и 3-х. Как следствие – лазеек для проникновения холода- гораздо больше.
Именно поэтому третью категорию рекомендуют применять при возведении нежилых построек.
Блоки третьей категории выглядят непрезентабельноСтеновые блоки могут иметь гладкую поверхность, либо обладать пазом и гребнем. В последнем случае, кладка стены значительно облегчится, а у новичка будет возможность выстроить практически идеальную поверхность.
Пеноблок выпускается также в виде изделий с офактуренной лицевой поверхностью. Приобретение таковых избавит застройщика от необходимости отделки здания снаружи.
Положительные и отрицательные стороны материала
Таблица 2. Плюсы пено- и газобетона:
| Значения плотности и прочности позволяют применять материал как в качестве утеплителя, так и при строительстве несущих конструкций. | |
Способность к теплосохранению | Обеспечивает сокращение затрат на утеплении зданий. |
Указывает на долговечность будущего строения и его атмосферную стойкость. | |
| Исключает вредное воздействие. Позволяет применять изделия при возведении специализированных учреждений. | |
| Также не менее важное преимущество. Возможность возгорания здания снижается. | |
Вариативность размеров, большое количество различных изготовителей |
|
| Изделия поддаются обработке сравнительно легко и это, согласитесь, удобно. | |
Особенно важный плюс, указывающий на возможность снизить затраты при возведении основания, так как устройство массивного фундамента, при строительстве легковесной конструкции вовсе не обязательно. | |
| Процесс строительства значительно ускорится за счет габаритов. | |
Возможность самостоятельного изготовления | Любой застройщик может попробовать себя в роли производителя. Это – не только полезный опыт, но и снижение затрат. |
| Группа материалов составляют высокую конкуренцию не только благодаря своим качествам, но и приемлемой цене. |
Таблица 2. Недостатки:
| К сожалению, данное свойство характеризует оба представителя группы. | |
| Несмотря на то, что у пенобетона данная способность снижена до 10-16% в связи с закрытой структурой пор, в защите он все равно нуждается. Для газобетона значение составляет вовсе 25%. | |
| Механических воздействий изделия боятся. | |
Покупка специального крепежа | Любые метизы не подойдут. Это обусловлено особенностями материала |
Слабая устойчивость к вырывающим нагрузкам | Требует планирования крепления тяжелых предметов еще на уровне проекта и их укрепления. |
Полезные советы при самостоятельном проведении кладочных работ
Поскольку все особенности материала мы уже знаем, давайте разбираться как же правильно его использовать так, чтобы данные свойства сохранились.
Подготовка инструментов и материала для кладки
Кладка начинается с подготовки инструментов и материала. Если решено применять клеевой состав, то его достаточно приобрести в магазине. Ассортимент достаточно широк.
Внимание стоит обратить на следующие показатели, указанные производителем:
- Жизнеспособность смеси;
- Пластичность;
- Теплопроводность;
- Устойчивость к перепадам температур;
- Прочность;
- Расход.
Усредненный расход составляет около 1,3-1,5 мешка смеси на 1 м3. Если же вы все-таки решили использовать раствор, то следует запастись цементом, водой, песком. Для замеса также понадобится бетономешалка.
Инструменты необходимы следующие:
Дрель с насадкой для смешиванияЭтапы возведения строения
Теперь – об этапах работ при кладке.
Таблица 4. Технология кладки: этапы работ:
| Необходимо гидроизолировать основание. Оно должно быть ровным, без видимых дефектов, таких как сколы, выпуклости. | |
| Кладка начинается с углов будущего здания. Причем первое изделие размещают в самой высокой его точке (углу, опять же). Натягивают нить, на которую в последующем ориентируются. | |
| Выкладывается первый ряд полностью, рекомендуется применять при этом раствор. | |
| Укладка производится со смещением шва, используется, как правило, клеевая смесь. Толщина шва должна быть минимально возможной. | |
| Контроль производят при помощи уровня, коррекция – при помощи киянки. Делать это специалисты рекомендуют максимально часто, так как жизнедеятельность раствора не позволяет долго ждать. Если клей схватится, изменить уже не удастся. | |
Необходимый этап при укладке. Рекомендуется производить армирование при укладке первого и каждого третьего-четвертого ряда. Рекомендуется производить армирование при укладке первого и каждого третьего-четвертого ряда. | |
| Применить можно ножовку, пилу или электроинструмент. | |
| По окончанию укладки стен, необходимо исполнить его устройство. |
Видео в этой статье: «Блоки стеновые из ячеистого бетона d600», продемонстрирует процесс кладки популярных изделий.
Заключение
Стеновые блоки из ячеистого бетона определенно заслуживают столь большой популярности. Это обосновано высокими значениями технических свойств, и достойными эксплуатационными показателями. Цена на изделия не менее конкурентная, а широкий ассортимент форм и размеров позволяет подобрать подходящие изделия любому потребителю.
beton-house.com
Характеристики ячеистых бетонных блоков
Главная|Блоки и перекрытия|Характеристики ячеистых бетонных блоков
Дата: 21 ноября 2016
Просмотров: 836
Коментариев: 0
Традиционно для строительства зданий использовались плиты, кирпич, древесина. Материалы обладали достоинствами, но не являлись взаимозаменяемыми. Возникла потребность разработать универсальный состав, обладающий увеличенной прочностью, повышенной звуко- и теплоизоляцией, легкообрабатываемый и экологичный. Так появились легкие блоки из ячеистого бетона. Они отличаются структурой, для которой характерны искусственно сформированные замкнутые полости. Размер ячеек 2-5 миллиметров. Полости занимают до 85% объема массива.
Материалы обладали достоинствами, но не являлись взаимозаменяемыми. Возникла потребность разработать универсальный состав, обладающий увеличенной прочностью, повышенной звуко- и теплоизоляцией, легкообрабатываемый и экологичный. Так появились легкие блоки из ячеистого бетона. Они отличаются структурой, для которой характерны искусственно сформированные замкнутые полости. Размер ячеек 2-5 миллиметров. Полости занимают до 85% объема массива.
Свойства
Блоки стеновые из ячеистого бетона отличаются следующими свойствами:
- повышенной прочностью;
- устойчивостью к колебаниям температуры;
Таблица сравнений характеристик ячеистых бетонов
- экологичностью;
- невосприимчивостью к образованию плесени;
- неподверженностью к гниению;
- увеличенным сроком эксплуатации.
Виды пористых составов
Известно 2 вида пористого бетона:
- Газобетон, полученный в результате перемешивания раствора цемента с порошком алюминия. При смешивании в результате взаимодействия образуются полости, пятикратно увеличивающие объем массива. Газобетон приобретает губчатую структуру. При помещении состава в автоклав смесь твердеет, изделия приобретают промышленный вид. Процесс возможен промышленным образом, требует специального оборудования.
- Пенобетон, произведенный путем добавления в цементный состав вспененных реагентов, образующих воздушные пузырьки. Пористость состав приобретает при тщательном смешивании, твердея, превращается в пенобетон. Специфика технологии предусматривает изготовление материала в условиях строительной площадки и кустарным образом.
 При смешивании в результате взаимодействия образуются полости, пятикратно увеличивающие объем массива. Газобетон приобретает губчатую структуру. При помещении состава в автоклав смесь твердеет, изделия приобретают промышленный вид. Процесс возможен промышленным образом, требует специального оборудования.
При смешивании в результате взаимодействия образуются полости, пятикратно увеличивающие объем массива. Газобетон приобретает губчатую структуру. При помещении состава в автоклав смесь твердеет, изделия приобретают промышленный вид. Процесс возможен промышленным образом, требует специального оборудования.Таблица свойств и характеристик пенобетона и газобетона
Разместите рядом разновидности изделий. Имеются визуальные отличия. Газобетонные изделия отличаются четкой геометрией, светлые, тонут. Пенобетон внешне аналогичен цементу, плавает на поверхности.
Назначение
Блоки из ячеистого бетона, в зависимости от концентрации пор, имеют разную сферу применения, условно разделяются на следующие типы:
- конструкционные составы, плотностью 600–1200 килограмм на метр кубический;
- теплоизоляционные композиты, с удельным весом 400–600 килограмм на метр кубический.

Пористость определяет плотность изделия. С увеличением объёма полостей возрастают звукоизоляционные и теплозащитные характеристики, но снижается прочность. Поэтому при строительстве капитальных наружных стен здания, для которых необходима повышенная прочность, используют более плотный пористый материал. Изделия конструкционного назначения применяют для сооружения:
- капитальных стен;
- внутренних малонагруженных стен;
- перегородок.
Ячеистые блоки хорошо подходят для строительства домов любой сложности
Мероприятия по строительной тепловой изоляции производят с помощью теплоизоляционных элементов, которые позволяют:
- утеплять поверхности железобетонных перекрытий;
- создавать теплоизоляционный контур многослойных конструкций стен;
- теплоизолировать перекрытия чердачных помещений;
- обеспечивать с помощью жаропрочных смесей тепловую изоляцию специального оборудования, поверхность которого нагревается до 700°С;
- защищать поверхности трубопроводов и технологического оборудования, рабочий температурный режим которого повышается до 400°С.

Многоэтажные здания, частные постройки из пористого бетона отличаются повышенными тепловыми свойствами по сравнению с кирпичными объектами.
Уменьшенные допуски размеров изделий позволяют осуществлять кладку с использованием специального клея с толщиной шва до 3 мм. Благодаря этому отсутствуют перемычки холода, образующиеся при кладке на цементный раствор.
Что выбрать: пенобетон или газобетон?
Разберемся, какие блоки ячеистого бетона лучше? Сложно однозначно ответить на вопрос. Ответ зависит от условий, где планируется использовать стеновой материал. Рассмотрим главные моменты:
- Газобетон целесообразно использовать для возведения несущих стен. Он обладает повышенной прочностью.
Таблица сравнения основных свойств пенобетона и газобетона
- По способности противостоять отрицательным температурам оба состава имеют равные свойства.
- Пенобетон менее гигроскопичен, превосходит газобетон по степени водопоглощения.
- Газобетон дороже, что связано с автоклавным методом производства.

Характеристики блоков из ячеистого бетона подтверждают экономическую целесообразность применения газобетона для возведения несущих конструкций. Вспененный состав используется для утепления и строительства перегородок помещений.
Достоинства пористого композита
Блоки из ячеистого бетона обладают множеством положительных свойств. Основные плюсы:
- Способность проводить тепло. По теплопроводности материал близок к древесине, но отсутствует ограничение на ширину возводимых стен. При толщине стен, построенных из композитов, соответствующих размерам кирпичной кладки, обеспечивается благоприятный тепловой режим помещения. Целостность, однородность конструкции здания обусловлена отсутствием потребности в специальных утеплителях. Это актуально для постройки частных объектов, где застройщик заинтересован экономить тепло и, естественно, материальные ресурсы.
- Экономичность. Незначительный вес пористого изделия, которое легче кирпича, позволяет минимизировать расходы на строительно-монтажные мероприятия и обустройство фундамента. Композит не требует дополнительного утепления, а применение специальных клеящих составов позволяет достичь экономии при кладке.
 Композит не требует дополнительного утепления, а применение специальных клеящих составов позволяет достичь экономии при кладке.
Композит не требует дополнительного утепления, а применение специальных клеящих составов позволяет достичь экономии при кладке.Достоинства ячеистого бетона
- Возможность пропускать насыщенный паром влажный воздух. С возрастанием коэффициента паропроницаемости улучшается микроклимат. Помещение из пористого бетона обеспечивает комфорт зимой и прохладу летом. Вентилируемость постройки снижает вероятность образования плесени, грибка.
- Устойчивость к воспламенению. Ячеистые блоки обладают высокой огнестойкостью, чем отличаются от древесины. Отпадает необходимость в дополнительной защите от воспламенения. Материал применяют как огнеупор.
- Увеличенная точность геометрических размеров. Допуски составляют ± 2 миллиметра, что позволяет выполнять минимальную толщину кладочного шва, уменьшить расход клеевой смеси, увеличить тепловую изоляцию стен.
Сравнение с кирпичом и древесиной
Положительные эксплуатационные характеристики блоков из ячеистого бетона позволяют успешно конкурировать с древесиной и камнем. Рассмотрим особенности материалов:
Рассмотрим особенности материалов:
- Обрабатываемость. В блок легко забиваются гвозди, он поддается обработке рубанком, быстро распиливается ножовкой.
- Масса. В отличие от тяжелого кирпича, ячеистые блоки более легкие. Это обеспечивает возможность использовать изделия увеличенных размеров и значительно сократить потребление материала для соединительных швов. Небольшой вес позволяет достичь экономии при транспортировке композитов на строительную площадку. Возводить постройки, используя пористые составы, удобно на грунтах, обладающих низкой несущей способностью и использовать для зданий легкие, менее массивные столбчатые фундаменты
Таблица сравнения теплопроводности ячеистого бетона с кирпичом и древесиной
- Воспламеняемость. Композит огнестоек, чем выгодно превосходит древесину.
- Способность проводить тепло. Благодаря высокой пористости лидирует вспененный состав, который обладает низкой теплопроводностью, и по своим характеристикам похож на древесину.
- Морозоустойчивость. Особенности структуры пористого бетона и кирпича позволяют им сохранять свои свойства на протяжении 100 циклов замораживания.
- Прочность. Пористый композит достаточно твердый, но менее прочен по сравнению с кирпичом, что связано со структурой. Приняв решение – возвести здание с перекрытием из тяжелых плит, применяйте для несущих конструкций кирпич, выдерживающий увеличенную нагрузку на единицу площади.
- Способность поглощать влагу. Коэффициент водопоглощения характеризует объем воды, который может впитать материал. Кирпич может поглощать влагу в объеме 8-12% массы. Он превосходит ячеистые составы, обладающие водопоглощением до 20%, а древесине с ними, вообще, сложно конкурировать.

Проанализировав параметры, сложно уверенно определить, что предпочтительнее использовать для строительства здания: пористые составы или обычный кирпич. Понятно, что у древесины область применения ограничена. Материалы имеют свои достоинства и недостатки, которые необходимо учитывать, выбирая сырье для постройки.
Размеры
Блоки стеновые из ячеистого бетона производятся на основе марок раствора М20 – M50, отличающегося прочностью на сжатие. При толщине 150-400 мм его ширина составляет 200-500 мм, а длина варьируется 400-600 мм.
Перегородчатые блоки изготавливаются из смеси марки М500. При постоянной ширине 400 мм, толщина составляет 100-150 мм, а длина –300-600 мм. Широкая номенклатура выпускаемых изделий позволяет применять элементы при решении строительных задач различного уровня сложности.
Особенности применения
Осуществляя постройку, установите пористые композиты рядом с кирпичной стеной, обеспечив между ними воздушный вентилируемый зазор 6 мм. При отсутствии вентилируемого пространства пары, проникая сквозь пористый состав, будут конденсироваться на кирпичной кладке, постепенно вызовут разрушение. Заполните пространство жестким утеплителем плотностью 90 килограмм на метр кубический. Выполните специальные отверстия для вентиляции.
Выполните специальные отверстия для вентиляции.
Изделия из ячеистого композита, как стеновые, так и перегородочные, характеризуются точностью геометрических параметров и высоким качеством. Здания, возведенные с использованием легких пористых материалов, отличаются высокими акустическими показателями, увеличенной комфортностью и высоким уровнем тепловой защиты.
pobetony.ru
Стеновые блоки и камни : Кирпич : Блоки из ячеистого бетона
СТЕНОВЫЕ МАТЕРИАЛЫ
| Энциклопедия |
КИРПИЧ
По составу и способу производства кирпич делится на две группы — керамический и силикатный.
- Керамический кирпич получают путем обжига глин и их смесей.
- Силикатный кирпич состоит примерно из 90% песка, 10% извести и небольшой доли добавок. Смесь отправляется в автоклав, а не в обжиговую печь, как в случае с керамическим кирпичом. На эти два материала существуют разные ГОСТы, и строители очень четко проводят между ними границу.
- Рядовой кирпич используется для внутренних рядов кладки или для внешних рядов, но с последующей штукатуркой. Рядовой кирпич может иметь на боковой стороне вдавленный геометрический рисунок (для лучшего сцепления со штукатурным раствором).
- Лицевой кирпич однородного цвета, имеет две гладкие, ровные лицевые поверхности (так называемые «тычок» и «ложок»). Он, как правило, пустотелый (то есть в его «теле» много пустот, что делает стену из такого кирпича «теплей»).

Печной кирпич бывает разных размеров. Иногда его делают с рельефом. Для топочной части каминов используют шамотный кирпич, изготовленный из огнеупорных глин и выдерживающий температуру выше 1000°С. Шамотный кирпич имеет песочно-желтый цвет и зернистую структуру.
Кирпич бывает:
- одинарный 250 х 120 х 65 мм.
- утолщенный, или полуторный 250 х 120 х 88 мм.
- двойной 250 х 120 х 138 мм (ГОСТ определяет его как камень керамический).
 По ГОСТу кирпич по цвету должен соответствовать образцу-эталону, утвержденному заводом.
По ГОСТу кирпич по цвету должен соответствовать образцу-эталону, утвержденному заводом.По способу формования кирпич бывает полусухого прессования и пластического.
Пластический способ формования наиболее распространенный в производстве полнотелого и пустотелого (эффективного) кирпича. При этом способе формования подготовленную глиняную массу влажностью 18 25 % направляют в приемный бункер ленточного пресса. При помощи шнека масса дополнительно перемешивается, уплотняется и выдавливается через выходное отверстие пресса. Непрерывно выходящий из пресса брус разрезают на отдельные части в соответствии с размерами изготавливаемых изделий автоматическое резательное устройство. Ленточные прессы снабжены вакуум-камерами, в которых из глиняной массы частично удалятся воздух. Вакуумирование массы повышает ее пластичность и уменьшает формовочную влажность, сокращает длительность сушки сырца и одновременно повышает его прочность.
Полусухим способом формования можно изготовлять кирпич из малопластичных глин, что расширяет сырьевую базу. Кроме того, существенное преимущество полусухого способа формования по сравнению с пластическим — применение глиняной массы с меньшей влажностью (8 12%), что значительно сокращает или даже исключает сушку сырца.
Кроме того, существенное преимущество полусухого способа формования по сравнению с пластическим — применение глиняной массы с меньшей влажностью (8 12%), что значительно сокращает или даже исключает сушку сырца.
При полусухом способе каждое изделие формуют отдельно на прессах, обеспечивающих двустороннее прессование под давлением 150 кгс/см2. Кирпич полусухого прессования имеет четкую форму, точные размеры, прочные углы и ребра.
По наличию пустот в теле кирпич лицевой и рядовой делится на пустотелый (эффективный) и полнотелый (камни керамические бывают только пустотелыми). Чем больше пустот (их может быть больше 50%), тем кирпич теплее.
Прочность кирпича основная характеристика способность материала сопротивляться внутренним напряжениям и деформациям, не разрушаясь.
Марка кирпича это показатель прочности, обозначается «М» с цифровым значением. Цифры показывают, какую нагрузку на 1 кв.см. может выдержать кирпич. Например, марка 100 (М100) обозначает, что кирпич гарантированно выдерживает нагрузку в 100 кг на 1 кв. см. Кирпич может иметь марку от 75 до 300. В продаже чаще всего встречается кирпич М100, 125, 150, 175.
см. Кирпич может иметь марку от 75 до 300. В продаже чаще всего встречается кирпич М100, 125, 150, 175.
Морозостойкость кирпича способность материала выдерживать попеременное замораживание и оттаивание в водонасыщенном состоянии. Морозостойкость (обозначается «Мрз») измеряется в циклах. Во время стандартных испытаний кирпич опускают в воду на 8 часов, потом помещают на 8 часов в морозильную камеру (это один цикл). И так до тех пор, пока кирпич не начнет менять свои характеристики (массу, прочность и т.п.). Тогда испытания останавливают и делают заключение о морозостойкости кирпича.
В соответствии с ГОСТом 530-95 «Кирпич и камни керамические. Технические требования» при транспортировке допускается бой не более 5%.
Условные обозначения кирпича:
- КР Стр рядовой полнотелый строительный
- КР Обл полнотелый облицовочный
- КРЭ Стр /Обл рядовой эффективный строит./облиц.
- КРЭУ Стр/Обл рядовой эффективный утолщенный
- КРУ полнотелый утолщенный
- КАЭ двойной эффективный
БЛОКИ ИЗ ЯЧЕИСТОГО БЕТОНА
Газосиликатный блок представляет собой искусственный камень с равномерно распределенными порами — ячейками. Пористая структура придает материалу ряд физико-механических свойств, которые делают
его эффективным строительным материалом.
Пористая структура придает материалу ряд физико-механических свойств, которые делают
его эффективным строительным материалом.
Главное отличие газосиликатных блоков от пенобетона в технологии производства. При изготовлении пенобетона, как и при изготовлении газосиликата, используются песок, цемент и порообразователь. Однако вспенивание производится не в результате химической реакции, а специальными миксерами. Полученная масса застывает естественным образом (без автоклавной обработки). Технология производства газосиликата требует более сложного и дорогостоящего оборудования. Тем не менее, при производстве пенобетона невозможно добиться такого сочетания прочности, плотности, теплопроводности, звукоизоляционных и других свойств, присущих газосиликату автоклавного твердения.
Важной характеристикой ячеистого бетона является его плотность. Чем плотность ниже (т.е. чем выше пористость), тем лучше теплозащита материала.
Блоки из ячеистого бетона дают самые теплые стены. Благодаря заключенному в порах воздуху теплоизоляционные свойства стен из ячеистого бетона в 3 5 раз выше, чем у кирпича и в 8 раз выше, чем у тяжелого бетона при одинаковых температурных условиях. Применение газосиликатных блоков в стенах малоэтажных домов взамен керамического кирпича позволяет сэкономить до 35% энергозатрат на отопление.
Благодаря заключенному в порах воздуху теплоизоляционные свойства стен из ячеистого бетона в 3 5 раз выше, чем у кирпича и в 8 раз выше, чем у тяжелого бетона при одинаковых температурных условиях. Применение газосиликатных блоков в стенах малоэтажных домов взамен керамического кирпича позволяет сэкономить до 35% энергозатрат на отопление.
Морозостойкость же блоков объясняется их мелкопористостью, обеспечивающей резервный объем, в который без разрушения материала вытесняется при замерзании расширяющиеся лед и вода. Считается, что материал морозостоек, если его водонасыщение не превышает критической величины 60% от массы. На практике же, при правильной эксплуатации, эта величина не превышает 35%.
Блоки из ячеистого бетона предназначены для кладки наружных, внутренних стен и перегородок зданий с относительной влажностью воздуха помещений не более 75% при неагрессивной среде. Из ячеистого бетона можно легко и быстро строить коттеджи. Загородный дом площадью 250 кв.м. пятью рабочими строится за 40 дней. Ячеистый бетон не гниет и не стареет.
пятью рабочими строится за 40 дней. Ячеистый бетон не гниет и не стареет.
Он легко обрабатывается: пилится, сверлится, гвоздится без образования трещин. В нем можно без особых усилий нарезать пазы, каналы и отверстия, например, для электропроводки, трубопроводов.
Материал достаточно легок. Стандартный блок размером 600х200х300 мм. марки D600 имеет массу 26 кг (после усыхания 22 кг) и может заменить в ограждающей стене 30 кирпичей, вес которых более 120 кг. Легкость позволяет снизить транспортные расходы и трудоемкость строительных работ. Процесс кладки ускоряется в 4 раза, а расход раствора уменьшается в 5 7 раз.
При низкой объемной массе газосиликат имеет довольно высокую прочность на сжатие. Максимальная этажность зданий с несущими станами составляет 4 этажа. При применении определенных конструктивных решений возможно использование блоков из ячеистого бетона в высотных зданиях без ограничения этажности.
Материал не горюч и абсолютно нетоксичен при пожаре. Ячеистый бетон «дышит», регулируя влажность в помещении, создавая оптимальный микроклимат.
Ячеистый бетон «дышит», регулируя влажность в помещении, создавая оптимальный микроклимат.
В соответствии с ГОСТом 21520-89 «Блоки из ячеистых бетонов стеновые мелкие. Технические требования» при транспортировке допускается бой не более 5%.
КАМНИ БЕТОННЫЕ СТЕНОВЫЕ
Камни (СКЦ) применяют в соответствии с действующими строительными нормами и правилами при возведении стен и других конструкций зданий и сооружений различного назначения.
В зависимости от назначения камни выпускают для кладки наружных и внутренних стен (порядовочные, угловые, перевязочные) и перегородок (перегородочные).
Камни изготавливают, как правило, в форме прямоугольного параллелепипеда. Номинальные размеры камней приведены в таблице 1.
Таблица 1
|
Торцы у камней могут быть плоскими, с пазами или иметь шпунт и гребень.
 Допускается изготавливать камни с одной плоской торцевой гранью.
Допускается изготавливать камни с одной плоской торцевой гранью.Углы у камней могут быть прямыми или закругленными.
Опорные поверхности камней могут быть плоскими или иметь продольные пазы, расположенные на расстоянии не менее 20 мм от боковой поверхности камня.
Камни изготавливают пустотелыми и полнотелыми. Масса камня должна быть не более 31 кг.
Пустоты необходимо располагать перпендикулярно опорной поверхности камня и распределять равномерно по его сечению. Пустоты могут быть сквозные и несквозные.
Толщина вертикальной диафрагмы (минимальная толщина перегородок) должна быть не менее 20 мм, горизонтальной диафрагмы для камней с несквозными пустотами не менее 10 мм.
Камни должны соответствовать требованиям стандарта и изготавливаться по технологическому регламенту, утвержденному предприятием-изготовителем.
На камне не допускаются дефекты внешнего вида, размеры и число которых превышают указанные в таблице 2.
Таблица 2
|
 , не более
, не болееЖировые или другие пятна размером более 10 мм. на лицевых поверхностях камней не допускаются.
Число камней с трещинами, пересекающими одно или два смежных ребра, а также количество половняка в партии должно быть не более 5%. Половняком считают изделие, состоящее из парных половинок или имеющее поперечную трещину, протяженностью их опорной поверхности более 1/2 ширины камня.
Стеновые керамзитобетонные блоки используются при возведении наружных стен и межкомнатных перегородок. Они применяются для заполнения каркаса при монолитном железобетонном домостроении, при строительстве хозяйственных построек, гаражей и коттеджей для индивидуального заказчика.
Применение керамзитобетона при возведении зданий и сооружений позволяет существенно усовершенствовать технологию и ускорить строительство.
Керамзитобетонные блоки имеют ряд преимуществ перед керамическим и силикатным кирпичом:
- удельный вес в 2. 5 раза ниже, чем при кирпичной кладке;
- содержание цемента в кладке из вибропрессованных блоков значительно ниже;
- один стандартный блок по объему соответствует семи кирпичам ;
- квалифицированный каменщик укладывает в смену объем стены в три раза больше, чем при кирпичной кладке.
 5 раза ниже, чем при кирпичной кладке;
5 раза ниже, чем при кирпичной кладке;Керамзитобетон «дышит», регулируя влажность воздуха в помещении. Строения из керамзитобетонных блоков вечны и не требуют ухода. Материал не гниет, не горит, в отличие от дерева, и не ржавеет, по сравнению с металлом, но обладает положительными свойствами дерева и камня одновременно.
Список литературы:
Шишкина Ирина © 
Цена блока из ячеистого бетона в Москве и обл.
Блоки стеновые из ячеистого бетона автоклавного твердения 1 категории – один из самых популярных видов материалов для строительства. Основная сфера применения блоков из ячеистого бетона – возведение наружных несущих стен малоэтажных домов (коттеджей, частных домов), а также строительство перегородок и самонесущих стен монолитно-каркасных зданий.
Блоки стеновые из ячеистого бетона автоклавного твердения выгодно сочетают в себе три качества: относительно небольшой вес, низкая цена блоков из ячеистого бетона и высокая прочность. Эти свойства позволяют при строительстве малоэтажных зданий отказаться от каркаса.
Наши цены:
Блоки из ячеистого бетона автоклавного твердения, первой категории (для кладки на клей) производства ОАО «Белорусский цементный завод», Д500 или Д600, упакованные в пленку на поддоне, с доставкой по городу Москва и области:
6270 руб/м3| Размер | Кол-во шт в 1 м3 | Цена за 1 м3 | Цена за 1 шт |
|---|---|---|---|
| 600x300x200 | 27. 7778 7778 | 6270 | 225.72 |
| 600х400х200 | 20.8333 | 6270 | 300.96 |
| 600x200x250 | 33.3333 | 6270 | 188.10 |
| 600x250x250 | 26.6666 | 6270 | 235.13 |
| 600x300x250 | 22.2222 | 6270 | 282.15 |
| 600x350x250 | 19.0476 | 6270 | 329.18 |
| 600x375x250 | 17.7936 | 6270 | 352.37 |
| 600x400x250 | 16.6666 | 6270 | 376.20 |
| 600x175x295 | 32.2841 | 6270 | 194.21 |
| 600x190x295 | 29. 7354 7354 | 6270 | 210.86 |
| 600x200x295 | 28.2485 | 6270 | 221.96 |
| 600x295x250 | 22.5988 | 6270 | 277.45 |
| 600x300x295 | 18.8235 | 6270 | 333.09 |
| 600x395x250 | 16.8776 | 6270 | 371.50 |
| 600x400x295 | 14.1243 | 6270 | 443.92 |
| 600x500x295 | 11.2994 | 6270 | 554.90 |
— Минимальный заказ — 1 машина (может комплектоваться разными размерами)
Блоки из ячеистого бетона автоклавного твердения, первой категории (для кладки на клей) производства ЧП ГазосиликатСтрой, Д500, упакованные в пленку на европоддоне, с доставкой по городу Москва и области:
6370 руб/м3| Размер | Кол-во шт в 1 м3 | Цена за 1 м3 | Цена за 1 шт |
|---|---|---|---|
| 200х300х600 | 27..jpg) 7778 7778 | 6370 | 229.32 |
| 250х200х600 | 33.3333 | 6370 | 191.10 |
| 250х300х600 | 22.2222 | 6370 | 286.65 |
| 250х400х600 | 16.6667 | 6370 | 382.20 |
Блоки из ячеистого бетона автоклавного твердения, первой категории (для кладки на клей) производства ООО Егорьевский Завод Строительных Материалов, Д500, упакованные в пленку на европоддоне, с доставкой по городу Москва и области:
6730 руб/м3| Размер | Кол-во шт в 1 м3 | Цена за 1 м3 | Цена за 1 шт |
|---|---|---|---|
| 250x200x625 | 32.0000 | 6370 | 199.06 |
| 250x250x625 | 25. 6000 6000 | 6370 | 248.83 |
| 250x300x625 | 21.3333 | 6370 | 298.59 |
| 250x350x625 | 18.2857 | 6370 | 348.36 |
| 250x375x625 | 17.0667 | 6370 | 373.24 |
| 250x400x625 | 16.0000 | 6370 | 398.13 |
| 250x500x625 | 12.8000 | 6370 | 497.66 |
| 200x500x625 | 16.0000 | 6370 | 398.13 |
— Минимальный заказ — 1 машина (может комплектоваться разными размерами)
— Цена указана с учетом поддонов и доставки в любую точку Москвы и Московской области.
Блоки из ячеистого бетона автоклавного твердения, первой категории (для кладки на клей) производства ОАО Могилевский КСИ РБ, Д500, упакованные в пленку на европоддоне, с доставкой по городу Москва и области:
6370 руб/м3| Размер | Кол-во шт в 1 м3 | Цена за 1 м3 | Цена за 1 шт |
|---|---|---|---|
| 200x290x590 | 29. 2227 2227 | 6370 | 217.98 |
| 200x300x600 | 27.7778 | 6370 | 229.32 |
| 200x400x600 | 20.8333 | 6370 | 305.76 |
| 200x500x625 | 16.0000 | 6370 | 398.13 |
| 250x175x625 | 36.5714 | 6370 | 174.18 |
| 250x200x625 | 32.0000 | 6370 | 199.06 |
| 250x250x625 | 25.6000 | 6370 | 248.83 |
| 250x300x625 | 21.3333 | 6370 | 298.59 |
| 250x350x625 | 18.2857 | 6370 | 348.36 |
| 250x375x625 | 17. 0667 0667 | 6370 | 373.24 |
| 250x400x625 | 16.0000 | 6370 | 398.13 |
| 250x500x625 | 12.8000 | 6370 | 497.66 |
— Минимальный заказ — 1 машина (может комплектоваться разными размерами)
Купить блоки можно в разделе Контакты или по телефону
Ячеистый бетон Хебель (Hebel) — статьи на тему Блоки строительные
Имея минимальный удельный вес, гарантирующий низкую теплопроводность (?теплые стены?), газобетон обладает высокой несущей способностью. Газобетон используется не только как несущий стеновой материал, но и как теплоизоляция.Автоклавный ячеистый бетон известен под разными именами: газобетон, газосиликат, газоблок ? обозначающие один и тот же по сути материал ? искусственный пористый камень.
Семейство ячеистых бетонов включает в себя и материалы, не проходящие автоклавную обработку. Они известны как пенобетон или пеноблоки и очень существенно отличаются по физико-техническим параметрам от бетонов, твердеющих в автоклавах.
Они известны как пенобетон или пеноблоки и очень существенно отличаются по физико-техническим параметрам от бетонов, твердеющих в автоклавах.
География применения автоклавного ячеистого бетона охватывает все климатические пояса и все континенты, Заводы по его производству расположены как в морозных Канаде и Сибири, так и в жарких Австралии, Южной и Северной Африке; автоклавный ячеистый бетон применяется в засушливой Аравийской пустыне и в муссонной Юго-Восточной Азии, в сейсмически активных Японии, Турции и Калифорнии, — одним словом ? везде.
За свою более чем восьмидесятилетнюю историю газобетонные блоки нашли применение практически во всех типах конструктивных элементов зданий и сооружений самого различного назначения. Этот универсальный материал используется для возведения несущих и ненесущих стен, а также для изготовления армированных плит перекрытий и покрытий.
Качество изделий из газоблоков напрямую зависит от используемого сырья, технологии изготовления и оборудования предприятия, и значительно отличается у разных производителей. Газобетонные Блоки, изготовленные в условиях автоматизированного заводского производства отличаются стабильно высокими качественными характеристиками ? точностью геометрических размеров, прочностью и плотностью. Ячеистый бетон производится на крупных заводах в Санкт-Петербурге, Эстонии и Латвии, Беларуси и на стройплощадку попадает в виде готовых блоков. Промышленные условия и используемые технологии при производстве ячеистого бетона обеспечивают блокам высочайшее качество и позволяют довести этот материал до совершенства по ряду параметров.
Газобетонные Блоки, изготовленные в условиях автоматизированного заводского производства отличаются стабильно высокими качественными характеристиками ? точностью геометрических размеров, прочностью и плотностью. Ячеистый бетон производится на крупных заводах в Санкт-Петербурге, Эстонии и Латвии, Беларуси и на стройплощадку попадает в виде готовых блоков. Промышленные условия и используемые технологии при производстве ячеистого бетона обеспечивают блокам высочайшее качество и позволяют довести этот материал до совершенства по ряду параметров.
Процесс производства ячеистого бетона напоминает выпекание хлеба: в смесителе замешивается вода, цемент, молотый кварцевый песок, тщательно размельченная известь, добавляется алюминиевая пудра в качестве газообразователя — и смесь ячеистого бетона готова. Использование высокотехнологичного резательного оборудования позволяет разрезать полученный массив с высокой точностью на блоки и плиты. В автоклавной печи ячеистый бетон твердеет под давлением в атмосфере насыщенного пара при температуре около 184 ºС. Образовавшаяся уникальная кристаллическая структура придает блокам его превосходные свойства. Применяемая технология производства обеспечивает равномерную плотность массива и наилучшие, среди ячеистых бетонов, показатели прочности. Микроструктуру газобетона определяют ‘открытые’ поры, нежели закрытые у пенобетона. Ячейки между собой имеют связи, поэтому при попадании воды и замерзании — не разрушаются. Испытания на заводе СИБИТ в Сибири доказали, что подобные блоки могут выдерживать 250 циклов заморозки/разморозки.
Образовавшаяся уникальная кристаллическая структура придает блокам его превосходные свойства. Применяемая технология производства обеспечивает равномерную плотность массива и наилучшие, среди ячеистых бетонов, показатели прочности. Микроструктуру газобетона определяют ‘открытые’ поры, нежели закрытые у пенобетона. Ячейки между собой имеют связи, поэтому при попадании воды и замерзании — не разрушаются. Испытания на заводе СИБИТ в Сибири доказали, что подобные блоки могут выдерживать 250 циклов заморозки/разморозки.
Безусловное достоинство стен из газобетонных блоков — высокая устойчивость к любым атмосферным воздействиям: дождю, снегу, сильным порывам ветра. Такой стене не страшны грибок, плесень и микроорганизмы, в других случаях предъставляющие серьезную угрозу для несущих конструкций. Стена из блоков механически устойчива и долговечна, хорошо переносит воздействие агрессивных сред и служит хорошей защитой от холода.
Легкость. Газобетон ? легкий строительный материал. Стандартный мелкий блок из ячеистого бетона (ГОСТ 21520-89), марки плотности Д500, размером 300х250х600 мм имеет массу 32 кг и может заменить окло 30-ти кирпичей, вес которых составляет 120 кг. Легкость газосиликата позволяет снизить транспортно-монтажные расходы и затраты на устройство фундаментов, снизить трудоемкость работ.
Легкость газосиликата позволяет снизить транспортно-монтажные расходы и затраты на устройство фундаментов, снизить трудоемкость работ.
Теплопроводность. Благодаря пористой структуре газосиликат является конструктивно- теплоизоляционным материалом. Стены, выполненные из газобетона, полностью соответствуют новым требованиям СНиП по сопротивлению теплопередаче ограждающих конструкций жилых и общественных зданий. Коэффициент теплопроводности газобетона в сухом состоянии ? 0,12 Вт/м C0 Заключенный в порах воздух приводит к исключительному теплоизоляционному эффекту. Термическое сопротивление ограждающих конструкций из ячеистого бетона в 3 раза выше, чем из керамического кирпича и в 8 раз выше, чем из тяжелого бетона. В процессе эксплуатации здания из ячеистого бетона расходы на отопление снижаются на 25-30 %.
Теплоаккумуляционные свойства газобетона Ячеистый бетон способен аккумулировать тепло. Он накапливает тепло от отопления или от солнечных лучей. При низких температурах, к примеру, ночью, когда отопление становится более слабым, отдает накопленное тепло во внутренние помещения. Вместе с высокой степенью теплоизоляции, а также благодаря аккумуляции тепла обеспечивается постоянная и комфортная температура во всем доме. Зимой происходит экономия топлива, а в летнее время сохраняется приятная прохлада. Применение этого материала позволяет значительно сэкономить на отоплении. По теплопроводности блоки стандартной толщины (375 мм) эквивалентны 600-миллиметровой кирпичной кладке.
Вместе с высокой степенью теплоизоляции, а также благодаря аккумуляции тепла обеспечивается постоянная и комфортная температура во всем доме. Зимой происходит экономия топлива, а в летнее время сохраняется приятная прохлада. Применение этого материала позволяет значительно сэкономить на отоплении. По теплопроводности блоки стандартной толщины (375 мм) эквивалентны 600-миллиметровой кирпичной кладке.
Звукоизоляционные свойства газобетона, благодаря его пористой ячеистой структуре, в 10 раз выше, чем у кирпичной кладки. В зданиях из ячеистого бетона обеспечиваются действующие требования по звукоизоляции.
Пожаробезопасность. Поскольку для изготовления газобетона берется лишь природное минеральное сырье, то нет и опасности возгорания. Газобетон, будучи неорганическим и негорючим материалом, выдерживает односторонне воздействие огня в течение 3-7 часов. Это ? материал, способный защитить металлические конструкции от прямого воздействия огня.
Морозостойкость. Газобетон морозостоек, что объясняется наличием резервных пор, в которые вытесняется при замерзании лед и вода. Сам материал при этом не разрушается. Считается, что при соблюдении технологии строительства морозостойкость материала не менее 25 циклов (максимум пока никто не оценивал).
Сам материал при этом не разрушается. Считается, что при соблюдении технологии строительства морозостойкость материала не менее 25 циклов (максимум пока никто не оценивал).
Прочность При плотности Д500(500 кг/м3) газобетон имеет высокую прочность на сжатие ? 28-40 кгс/см3, и класс бетона по прочности В2(2,5) за счет автоклавной обработки. Материал может использоваться для кладки несущих стен, стенового заполнения каркасных высотных зданий, а также для кладки внутренних стен и перегородок.
Экономичность и быстрота возводимых конструкций За счет относительно больших габаритов газобетонного блока и его малого веса (не требует специальных подъемных механизмов) существенно (по некоторым оценкам ? в 4 раза) возрастает скорость строительства. Соответственно меньше становятся трудозатраты. Вместо стандартного раствора используется клеевой. Все это уменьшает общую стоимость возводимого здания. В процессе эксплуатации здания из ячеистого бетона расходы на отопление снижаются на 25-30 %.
Конструкционность. Точные геометрические характеристики изделий позволяют вести кладку блоков с использование клеевого раствора, который обеспечивает прочность сцепления и исключает наличие в кладке ?мостиков холода?.
Простота обработки. Газобетон легко обрабатывается любым режущим инструментом, пилится, сверлится, гвоздится, строгается, штрабится ? это делает его применение особенно привлекательным. Простота обработки ячеистого бетона позволяет создавать интересные архитектурные решения, в том числе прорезать каналы и отверстия под розетку, электропроводку, трубопроводы, арочные конфигурации.
Экологичность Современный газобетон производится из песка, извести, цемента и алюминиевой пудры. Он не выделяет токсичных веществ и по своей экологичности уступает лишь дереву. Но при этом газобетон, в отличие от дерева, не гниет и не стареет. Экологическая чистота применяемых сырьевых материалов гарантирует полную безопасность газобетонных изделий для человека. Радиационный фон газобетона не превышает 9-11 мкр/ч. Это пористый материал, поэтому в доме, построенном из газобетона, дышится так же легко, как и в деревянном. Производство ячеистого бетона не требует больших затрат энергии, т.к. бетон затвердевает под воздействием пара при температуре всего лишь 180С. Вторичное применение отработавшего пара и вторичная переработка обеспечивают возврат энергии и воды в производственном кругообороте. Отходы производства могут быть применены вторично, что также является вкладом в дело охраны окружающей среды.
Это пористый материал, поэтому в доме, построенном из газобетона, дышится так же легко, как и в деревянном. Производство ячеистого бетона не требует больших затрат энергии, т.к. бетон затвердевает под воздействием пара при температуре всего лишь 180С. Вторичное применение отработавшего пара и вторичная переработка обеспечивают возврат энергии и воды в производственном кругообороте. Отходы производства могут быть применены вторично, что также является вкладом в дело охраны окружающей среды.
Ячеистый бетон обладает свойствами, с одной стороны, камня, с другой — дерева. Сочетание этих свойств делает его прекрасным строительным материалом. Ячеистый бетон обладает прочностью камня. Один блок или панель выдерживают сжатие, измеряемое несколькими десятками тонн. Как камень он несгораем, а также практически не реагирует с водой или влагой. С другой стороны, ячеистый бетон обладает легкостью и обрабатываемостью, свойственными дереву. Как и дерево, он пилится, сверлится, строгается и фрезеруется. В нем легко можно устраивать различные крепления и выполнять проводку. Благодаря тому, что до 80% объема ячеистого бетона заполнено воздушными ячейками, он обладает прекрасной теплоизоляционной способностью, т.е. используя ячеистый бетон, другие теплоизоляционные материалы не нужны.
В нем легко можно устраивать различные крепления и выполнять проводку. Благодаря тому, что до 80% объема ячеистого бетона заполнено воздушными ячейками, он обладает прекрасной теплоизоляционной способностью, т.е. используя ячеистый бетон, другие теплоизоляционные материалы не нужны.
Дома из ячеистого бетона в различных климатических условиях практически вечные и не требуют особого ухода. Материал не гниет, в отличие от дерева, и не ржавеет по сравнению с металлом. Здание, построенное из ячеистого бетона, не сгорает, его конструкции остаются в неизменном виде и для устранения последствий пожара требуется лишь обновление поверхностных покрытий и внутренней отделки. В то же время при пожаре деревянный дом сгорает дотла, и даже стальные конструкции перекашиваются и рушатся.
Из ячеистого бетона строят жилые дома, промышленные и общественные здания, а также различные сельхозпостройки. Простота конструкции из ячеистого бетона и его высокие теплоизоляционные свойства получили положительные оценки при использовании, как в теплых, так и в холодных климатических условиях. Массивность материала является свойством, выравнивающим внутри здания температурные колебания, как в летнюю жару, так и в зимний мороз. Величина коэффициента теплопроводности ячеистого бетона удовлетворяют жестким требованиям, предъявляемым при ведении строительства в различных регионах земного шара. Ячеистый бетон обладает относительно высокой морозостойкостью.
Массивность материала является свойством, выравнивающим внутри здания температурные колебания, как в летнюю жару, так и в зимний мороз. Величина коэффициента теплопроводности ячеистого бетона удовлетворяют жестким требованиям, предъявляемым при ведении строительства в различных регионах земного шара. Ячеистый бетон обладает относительно высокой морозостойкостью.
Термическое сопротивление ограждающих конструкций из ячеистого бетона, при прочих равных условиях, в три раза выше, чем у глиняного кирпича и в восемь раз выше, чем у тяжелого бетона. Особенностью домов из ячеистого бетона является то, что они не имеют ‘мостиков холода’. В процессе эксплуатации здание из ячеистого бетона позволяет снизить на 25-30 % расходы на отопление.
Ячеистый бетон обладает высокой способностью к поглощению звука. У зданий из ячеистого бетона обычно обеспечиваются действующие требования по звукоизоляции без каких-либо дополнительных мероприятий.
Для защиты стеновых конструкций из ячеистого бетона от климатических воздействий достаточно обработать стену любым современным гидрофобизирующим составом, наружные стены можно облицовывать кирпичом (с воздушной прослокой), диким камнем, сайдингом, фасадными системами или слоем штукатурки.
Ячеистый бетон — экологически чистый строительный материал. Согласно исследованиям, проведенным в Германии институтом лучевой гигиены Федеративного Управления по здравоохранению, уровень радиоактивности ячеистого бетона значительно ниже допустимых пределов. Многочисленные испытания показали, что содержание естественных радионуклидов в ячеистом бетоне не превышает 35 Бер. Кроме того, ячеистый бетон не выделяет токсичных веществ и газов.
Любые блоки по сути являются одним из наиболее быстрых и экономически целесообразных способов возведения стен, а блоки из ячеистого бетона являются, бесспорно, самым простым решением кладки стен зданий: жилых домов, сельскохозяйственных строений и небольших построек промышленного и складского назначения. Использование блоков не накладывает никаких ограничений на планировку зданий, его форму или высоту — из блоков может быть построено здание практически любого типа. При ведении строительства с применением блоков из ячеистого бетона на рабочей площадке не требуется специального подъемного оборудования. Блоки укладываются наподобие кирпичной кладки и связываются традиционным строительным раствором или же специальным клеем для ячеистых блоков. Толщина слоя клея составляет всего 2-3 мм; в таких швах, естественно, не образуются ‘мостики холода’.
Блоки укладываются наподобие кирпичной кладки и связываются традиционным строительным раствором или же специальным клеем для ячеистых блоков. Толщина слоя клея составляет всего 2-3 мм; в таких швах, естественно, не образуются ‘мостики холода’.
Наши строительные коллективы ещ? 15 лет назад отказались от строительства из других стеновых материалов — кирпич, сборный бетон и т.д. И сделали правильный выбор — материал приятный как в работе, так и прекрасен по тепло-физическим свойствам.
Плазматическая мембрана . Мембрана, окружающая клетку, состоит из двух липидных слоев, называемых «билипидной» мембраной. Липиды, присутствующие в плазматической мембране, называются «фосфолипидами». Эти липидные слои состоят из ряда строительных блоков жирных кислот. Жирная кислота, из которой состоит эта мембрана, имеет две разные части: небольшую водолюбивую головку и гидрофильную головку. Этот хвост гидрофобный. Hydro означает воду, а phobic означает страх. Плазматическая мембрана устроена таким образом, что хвосты обращены друг к другу с внутренней стороны, а головки обращены к внешней стороне мембраны. | ||
Каналы/поры — Каналы в клеточной плазматической мембране. Этот канал состоит из определенных белков, которые контролируют движение молекул, включая пищу и воду, в клетку. | ||
Клеточная стенка и плазмодесмы — В дополнение к клеточным мембранам растения имеют клеточные стенки. Клеточные стенки обеспечивают защиту и поддержку растений. У наземных растений клеточная стенка в основном состоит из целлюлозы. В отличие от клеточных мембран, материалы не могут проникать сквозь клеточные стенки. Эти отверстия используются для сообщения и транспортировки материалов между растительными клетками, поскольку клеточные мембраны могут соприкасаться и, следовательно, обмениваться необходимыми материалами. | ||
Перегородки и поры клеточной стенки — Грибковые клетки имеют как клеточные мембраны, так и клеточные стенки, как клетки растений. Клеточные стенки обеспечивают защиту и поддержку. Клеточные стенки грибов в основном состоят из хитина, того же вещества, что и экзоскелеты насекомых. Поскольку материалы не могут проникнуть сквозь клеточные стенки, клетки грибов имеют специальные отверстия, называемые порами. Материалы могут перемещаться между грибковыми клетками через поры. Некоторые грибковые клетки также имеют перегородки (множественное число — перегородки), представляющие собой специальные внутренние стенки между клетками, которые находятся в длинных трубчатых нитях или тяжах, называемых гифами. | ||
| Клеточная капсула — Бактериальные клетки имеют клеточную мембрану и клеточную стенку, а также клеточную капсулу. Этот внешний слой часто состоит из сахаров или специальных белков. Это помогает защитить бактерии от поедания более крупными клетками, такими как иммунные клетки животных, и от заражения вирусами. вернуться к началу | ||
| Ядро — Ядро является центром управления клетки. Это самая большая органелла в клетке, содержащая ДНК клетки. ДНК и ядрышки ДНК (дезоксирибонуклеиновая кислота) содержит всю информацию, необходимую клеткам для жизни, выполнения своих функций и размножения. Внутри ядра находится еще одна органелла, называемая ядрышком . Ядрышко отвечает за образование рибосом. Кружки на поверхности ядра – это ядерные поры.Это места, где рибосомы и другие материалы перемещаются в ядро и из него. | ||
Нуклеоид — Бактерии не имеют ядра для хранения своей ядерной ДНК. | ||
| Плазмида — Помимо нуклеоида, бактерии имеют плазмиды.Плазмиды — это небольшие молекулы ДНК, которые могут содержать дополнительные гены, которые можно использовать, когда клетка находится в определенных условиях. Эти маленькие стопки ДНК также могут обмениваться между бактериальными клетками. вернуться к началу | ||
Эндоплазматический ретикулум (ЭР) — Это сеть мембран по всей цитоплазме клетки. Существует два типа ЭР. Когда рибосомы прикреплены, это называется шероховатым ЭР и гладким ЭР, когда рибосомы не прикреплены. Шероховатая эндоплазматическая сеть – это место, где в клетке происходит большая часть синтеза белка. Функция гладкой эндоплазматической сети заключается в синтезе липидов в клетке. | ||
| Рибосомы — Органеллы, участвующие в синтезе белков. Рибосомы состоят из двух частей, называемых субъединицами.
Они получили свое название из-за своего размера.Одна единица больше другой, поэтому их называют большими и малыми субъединицами. Обе эти субъединицы необходимы для синтеза белка в клетке. Когда две единицы стыкуются вместе со специальной информационной единицей, называемой матричной РНК, они образуют белки. Некоторые рибосомы находятся в цитоплазме, но большинство из них прикреплено к эндоплазматическому ретикулуму. Будучи прикрепленными к ER, рибосомы производят белки, которые нужны клетке, а также те, которые должны быть экспортированы из клетки для работы в других частях тела. | |
Комплекс Гольджи — Это органелла в клетке, которая отвечает за сортировку и правильную доставку белков, произведенных в ЭПР. Эта органелла была названа в честь итальянского врача Камилло Гольджи . Он был первым, кто описал эту органеллу в клетке. Это также единственная органелла с заглавной буквы. | ||
Митохондрия — Это электростанция клетки. Эта органелла упаковывает энергию из пищи, которую вы едите, в молекулы АТФ. Каждый тип клеток имеет разное количество митохондрий (множественное число).В клетках, которые должны выполнять большую работу, больше митохондрий, например, в клетках мышц ног, клетках сердечной мышцы и т. д. Другим клеткам требуется меньше энергии для выполнения своей работы и у них меньше митохондрий. | ||
Хлоропласт — Это органелла, в которой происходит фотосинтез. В этой органелле световая энергия солнца преобразуется в химическую энергию. Хлоропласты встречаются только в клетках растений, а не в клетках животных.Химическая энергия, вырабатываемая хлоропластами, в конечном итоге используется для производства углеводов, таких как крахмал, которые хранятся в растении. Хлоропласты содержат крошечные пигменты, называемые хлорофиллами . Хлорофиллы отвечают за улавливание световой энергии солнца. | ||
Везикулы — Этот термин буквально означает «маленький сосуд». Эта органелла помогает хранить и транспортировать продукты, вырабатываемые клеткой. Везикулы — это средства транспортировки и доставки, такие как наша почта и грузовики Federal Express. Некоторые везикулы доставляют материалы в части клетки, а другие транспортируют материалы за пределы клетки в процессе, называемом экзоцитозом. | ||
Пероксисомы — Они собирают и безопасно расщепляют химические вещества, токсичные для клетки. | ||
Лизосомы . Созданные аппаратом Гольджи, они помогают расщеплять большие молекулы на более мелкие части, которые клетка может использовать. | ||
Вакуоль — Растительные клетки имеют что-то похожее на очень большое пустое пространство посередине. Это пространство называется вакуолью. Не дайте себя обмануть, вакуоль содержит большое количество воды и хранит другие важные материалы, такие как сахара, ионы и пигменты. | ||
Центриол | ||
MicroTubule-Organizate Center (MTOC) — Fungi имеют MTOC, отличную от центриолей, найденных в большинстве. | ||
Микротрубочки — Трубчатые структуры, поддерживающие клетки. Микротрубочки можно найти в клетках любого животного, растения или гриба. Часть цитоскелета, обнаруженная в клетках животных, растений и грибов. Некоторые бактерии также имеют микротрубочки, но не все бактерии. | ||
Шпитценкерпер — Центр роста трубчатых грибковых клеток.Шпитценкорпер состоит из множества мелких пузырьков и плотных микрофиламентов. | ||
Актиновые филаменты — Длинные нити из более мелких единиц, которые играют важную роль в клеточной структуре. Участвует в изменении формы клеток во многих видах клеток. Часть цитоскелета, обнаруженная в клетках животных, растений и грибов.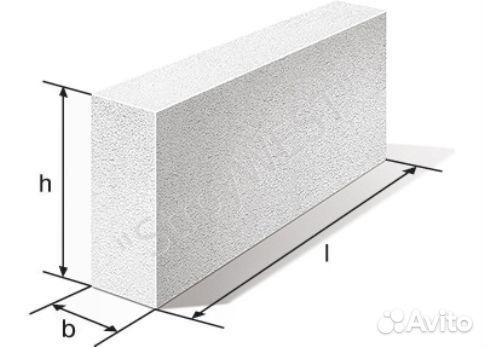 вернуться к началу | ||
| Цитоскелет — Состоит из нитей и канальцев, помогает формировать и поддерживать клетку.Это также помогает вещам перемещаться в камере. В художественных целях цитоскелет показан только в одном месте животной клетки, тогда как в действительности он встречается во всей клетке. вернуться к началу | ||
Цитоплазма — Термин для всего содержимого клетки, кроме ядра. Несмотря на то, что мультяшные рисунки не похожи на это, цитоплазма содержит в основном воду. Несколько забавных фактов о воде и человеческом теле:
наверх | ||
Бактериальные пили — Длинные нитевидные нити, отходящие от поверхности клетки. | ||
| Жгутик — хвост, прикрепленный к основному телу клетки, который может вращаться для перемещения клетки вперед.Чаще всего ассоциируется с бактериальными клетками. вернуться к началу |
 Hydro означает воду, а philic означает симпатию или любовь.Другая часть этой жирной кислоты представляет собой длинный водоотталкивающий или водоненавидящий хвост.
Hydro означает воду, а philic означает симпатию или любовь.Другая часть этой жирной кислоты представляет собой длинный водоотталкивающий или водоненавидящий хвост. Это было бы проблемой для растительных клеток, если бы не специальные отверстия, называемые плазмодесмами.
Это было бы проблемой для растительных клеток, если бы не специальные отверстия, называемые плазмодесмами.
 Вместо этого их ДНК находится в нуклеоиде. Эта структура не имеет защитной мембраны, но представляет собой плотно упакованный материал ДНК, а также содержит некоторое количество РНК и белков.
Вместо этого их ДНК находится в нуклеоиде. Эта структура не имеет защитной мембраны, но представляет собой плотно упакованный материал ДНК, а также содержит некоторое количество РНК и белков.  Гладкий ER также помогает в детоксикации вредных веществ в клетке.
Гладкий ER также помогает в детоксикации вредных веществ в клетке.  Точно так же, как наши почтовые посылки, которые должны иметь правильный адрес доставки, белки, произведенные в ER, должны быть правильно отправлены по соответствующему адресу. Это очень важный этап синтеза белка. Если комплекс Гольджи ошибается в доставке белков по нужному адресу, определенные функции в клетке могут прекратиться.
Точно так же, как наши почтовые посылки, которые должны иметь правильный адрес доставки, белки, произведенные в ER, должны быть правильно отправлены по соответствующему адресу. Это очень важный этап синтеза белка. Если комплекс Гольджи ошибается в доставке белков по нужному адресу, определенные функции в клетке могут прекратиться.

 клетки животных, но он выполняет аналогичную работу.MTOC строит микротрубочки, которые помогают построить внутреннюю клеточную структуру для формы и поддержки.
клетки животных, но он выполняет аналогичную работу.MTOC строит микротрубочки, которые помогают построить внутреннюю клеточную структуру для формы и поддержки.  Бактерии могут использовать их для связывания с другими бактериальными клетками для обмена генетическим материалом.
Бактерии могут использовать их для связывания с другими бактериальными клетками для обмена генетическим материалом. Новый строительный блок в строительстве стен растений — ScienceDaily
Исследователи из Университета Аделаиды в составе междисциплинарной международной группы открыли новый биохимический механизм, лежащий в основе жизни растений.
Исследование, опубликованное в The Plant Journal , подробно описывает открытие ферментативной реакции с участием углеводов, присутствующих в стенках клеток растений, которые необходимы для их структуры.
Руководитель проекта, профессор Мария Хрмова, сказала, что это открытие способствует получению важных знаний о том, как могут формироваться, структурироваться и реконструироваться клеточные стенки растений.
«Стены клеток растений выполняют ряд важных функций, в том числе придают форму множеству различных типов клеток, необходимых для формирования тканей и органов растения, межклеточной коммуникации, и они играют роль во взаимодействиях растений и микробов, включая защитные реакции против потенциальных патогенов», — сказала профессор Хрмова.
Более ранние исследования химии и функции ксилоглюкановых углеводов в растениях показали, что ферменты ксилоглюкановой ксилоглюкозилтрансферазы являются одним из ключевых ускорителей ремоделирования клеточных стенок.
Только благодаря развитию методологии, используемой в этом исследовании, рекомбинантной технологии, которая позволяет выделять белки в чистом виде, и доступности определенных углеводов стало возможным наблюдать ферментативную реакцию. который происходит между ксилоглюканом и пектиновыми углеводами.
«Когда мы смогли внимательно изучить субстратную специфичность ксилоглюкановых ксилоглюкозилтрансфераз ячменя, мы обнаружили химическую реакцию, которая приводит к образованию гетерополисахарида (углевода, состоящего из химически различных компонентов). Мы также смогли изучить эти реакции. на молекулярном уровне, чтобы определить, как именно работают эти ферменты», — сказала профессор Хрмова.
Мы также смогли изучить эти реакции. на молекулярном уровне, чтобы определить, как именно работают эти ферменты», — сказала профессор Хрмова.
«Одно дело — уметь идентифицировать различные компоненты клеточных стенок растений, но этого недостаточно, нам нужно понять, как они образуются и что они делают, и этот метод выделения чистых белков, чтобы их можно было осмотрели, позволили нам это сделать», — сказала профессор Хрмова.
«Это открытие является новым строительным блоком в нашем понимании того, как может быть построена клеточная стенка.»
«После того, как вы поймете, как что-то делается, вы сможете по-разному его конструировать или деконструировать, — говорит профессор Хрмова.
«Вот почему так ценны фундаментальные знания о том, как функционируют эти ферменты.»
Полученные результаты могут иметь далеко идущие последствия для устойчивости отраслей, основанных на растениеводстве, таких как сельское хозяйство, садоводство, лесное хозяйство для производства биотоплива и переработки продуктов питания и материалов.
На сегодняшний день команда охарактеризовала четыре из 36 ксилоглюкановых ксилоглюкозилтрансфераз в ячмене, так что еще многое предстоит изучить, что может привести к дальнейшим открытиям. Как только эта работа будет завершена для ячменя, методология может быть применена для изучения клеточных стенок других культур, таких как пшеница и рис.
«Растения являются крупнейшим возобновляемым ресурсом в мире — растения кормят мир, а также производят энергию в виде биотоплива», — сказала профессор Хрмова.
Эти знания могут помочь в биоинженерии подобных белков, участвующих в ремоделировании клеточных стенок растений, для создания продуктов более высокого качества и изучения способов деконструкции клеточных стенок растений для получения биотоплива.
Источник истории:
Материалы предоставлены Университетом Аделаиды . Оригинал написан Келли Браун. Примечание. Содержимое можно редактировать по стилю и длине.
Границы | Клеточная стенка растений: сложная и динамичная структура, выявленная ответами генов в условиях стресса
Введение
Клеточная стенка растений представляет собой сложную структуру, выполняющую разнообразные функции на протяжении всего жизненного цикла растения. В дополнение к поддержанию структурной целостности за счет сопротивления внутреннему гидростатическому давлению клеточная стенка обеспечивает гибкость для поддержки клеточного деления, биохимический каркас, обеспечивающий дифференцировку, а также патологический и экологический барьер, защищающий от стресса (Scheller and Ulvskov, 2010; Hamann, 2012; Такер и Колтунов, 2014). Клеточная стенка содержит широкий спектр рецепторов, пор и каналов, которые регулируют движение молекул и ответы на локальные и дальнодействующие элиситоры, включая гормоны, сахара, белки и РНК.В соответствии с ролью во многих процессах, структура клеточной стенки растений невероятно разнообразна не только между видами растений, но и между типами тканей. В общем, два типа стенок, окружающих растительные клетки, часто называют первичной стенкой и вторичной стенкой. Динамическая первичная стенка устанавливается в молодых клетках во время деления и обеспечивает гибкость и базовую структурную поддержку, защищая клетку и опосредуя межклеточные взаимодействия.
В дополнение к поддержанию структурной целостности за счет сопротивления внутреннему гидростатическому давлению клеточная стенка обеспечивает гибкость для поддержки клеточного деления, биохимический каркас, обеспечивающий дифференцировку, а также патологический и экологический барьер, защищающий от стресса (Scheller and Ulvskov, 2010; Hamann, 2012; Такер и Колтунов, 2014). Клеточная стенка содержит широкий спектр рецепторов, пор и каналов, которые регулируют движение молекул и ответы на локальные и дальнодействующие элиситоры, включая гормоны, сахара, белки и РНК.В соответствии с ролью во многих процессах, структура клеточной стенки растений невероятно разнообразна не только между видами растений, но и между типами тканей. В общем, два типа стенок, окружающих растительные клетки, часто называют первичной стенкой и вторичной стенкой. Динамическая первичная стенка устанавливается в молодых клетках во время деления и обеспечивает гибкость и базовую структурную поддержку, защищая клетку и опосредуя межклеточные взаимодействия. Более толстая и прочная вторичная стенка находится между первичной стенкой и плазматической мембраной и откладывается на более поздней стадии, когда клетка перестает расти и делиться.Вторичная стенка рассматривается как важная адаптация, которая позволяет наземным растениям выдерживать и способствовать вертикальному росту.
Более толстая и прочная вторичная стенка находится между первичной стенкой и плазматической мембраной и откладывается на более поздней стадии, когда клетка перестает расти и делиться.Вторичная стенка рассматривается как важная адаптация, которая позволяет наземным растениям выдерживать и способствовать вертикальному росту.
Типичные компоненты клеточной стенки включают целлюлозу, нецеллюлозные и пектиновые полисахариды, белки, фенольные соединения и воду. Основными компонентами (> 90%) являются полисахариды, структура и биосинтез которых были подробно рассмотрены в последнее время (Atmodjo et al., 2013; Pauly et al., 2013; Rennie and Scheller, 2014; Kumar et al., 2016). Вкратце, целлюлоза представляет собой нерастворимый в воде углевод, присутствующий как в первичных, так и во вторичных клеточных стенках, волокнистая структура которых позволяет поддерживать структурную целостность. Пектины, которые, возможно, являются наиболее сложными и гетерогенными полисахаридами клеточной стенки, существуют преимущественно в первичной клеточной стенке и играют роль в расширении, прочности, пористости, адгезии и межклеточной передаче сигналов. Другие распространенные нецеллюлозные полисахариды включают ксилоглюкан, β-1,3:1,4-глюкан, ксилан, маннан и каллозу, которые выполняют различные роли в механической поддержке, запасании и развитии.В отличие от целлюлозы пектиновые и нецеллюлозные полисахариды можно дополнительно отличить по замещению сахара и боковым цепям, которые присоединяются к полисахаридному остову во время биосинтеза (Шеллер и Ульвсков, 2010). Эти заместители влияют на растворимость, вязкость и взаимодействие с другими полисахаридами и белками в клеточной стенке.
Другие распространенные нецеллюлозные полисахариды включают ксилоглюкан, β-1,3:1,4-глюкан, ксилан, маннан и каллозу, которые выполняют различные роли в механической поддержке, запасании и развитии.В отличие от целлюлозы пектиновые и нецеллюлозные полисахариды можно дополнительно отличить по замещению сахара и боковым цепям, которые присоединяются к полисахаридному остову во время биосинтеза (Шеллер и Ульвсков, 2010). Эти заместители влияют на растворимость, вязкость и взаимодействие с другими полисахаридами и белками в клеточной стенке.
Функция различных компонентов клеточной стенки и то, как они взаимодействуют с экзогенными стимулами, такими как патогены и стресс окружающей среды, вызывает интерес в течение многих лет, особенно в связи с поиском механизмов, с помощью которых можно повысить устойчивость к патогенам, стрессоустойчивость и повысить урожайность сельскохозяйственных культур. достигнуто.В недавних обзорах изучалось, как абиотические сигналы изменяют биосинтез целлюлозы (Wang et al. , 2016), как экспансины и пероксидазы влияют на жесткость стенок во время стресса (Tenhaken, 2014) и как модификации нецеллюлозных полисахаридов, таких как ксилоглюкан, сопровождают стрессовые реакции (Le Галл и др., 2015). Только недавно в исследованиях начали рассматривать более широкое представление о том, как различные стрессы могут индуцировать сходные изменения в обилии транскриптов (Coolen et al., 2016). Вопрос о том, реагируют ли компоненты клеточной стенки, семейства генов, связанных с клеточной стенкой, или отдельные ортологичные гены клеточной стенки одинаковым образом на различные стрессы у разных видов, подробно не рассматривался, но может дать новые мишени с широкой специфичностью для модификация стрессовых реакций.Некоторые ответы могут быть скрыты в общедоступных наборах данных транскриптомов однодольных и двудольных видов, в которых подробно описаны глобальные транскрипционные реакции на патогены, такие как бактерии, грибы, оомицеты, насекомые и нематоды, а также на абиотические стрессы, такие как засуха, холод и жара.
, 2016), как экспансины и пероксидазы влияют на жесткость стенок во время стресса (Tenhaken, 2014) и как модификации нецеллюлозных полисахаридов, таких как ксилоглюкан, сопровождают стрессовые реакции (Le Галл и др., 2015). Только недавно в исследованиях начали рассматривать более широкое представление о том, как различные стрессы могут индуцировать сходные изменения в обилии транскриптов (Coolen et al., 2016). Вопрос о том, реагируют ли компоненты клеточной стенки, семейства генов, связанных с клеточной стенкой, или отдельные ортологичные гены клеточной стенки одинаковым образом на различные стрессы у разных видов, подробно не рассматривался, но может дать новые мишени с широкой специфичностью для модификация стрессовых реакций.Некоторые ответы могут быть скрыты в общедоступных наборах данных транскриптомов однодольных и двудольных видов, в которых подробно описаны глобальные транскрипционные реакции на патогены, такие как бактерии, грибы, оомицеты, насекомые и нематоды, а также на абиотические стрессы, такие как засуха, холод и жара. Эти наборы данных предоставляют ресурсы для идентификации семейств генов, связанных с углеводами, которые кодируют белки с аналогичными функциональными доменами База данных углеводно-активных ферментов (CAZy); (Lombard et al., 2014), а также определить, могут ли определенные семейства, такие как гликозилтрансферазы (GTs), гликозилгидролазы (GHs) и другие ферменты, модифицирующие углеводы, играть ключевую роль в синтезе и модификации клеточной стенки во время стресса.Поэтому после первых двух разделов этого обзора, где мы рассмотрели новые и исторические данные о роли полимеров и генов, связанных с клеточной стенкой, во время биотического и абиотического стресса, мы пересмотрели несколько наборов транскрипционных данных, чтобы обобщить реакцию клеточной стенки и гены и семейства, связанные с углеводами, при стрессе, подчеркивая замечательный уровень сохранения реакций, вызванных различными типами стресса.
Эти наборы данных предоставляют ресурсы для идентификации семейств генов, связанных с углеводами, которые кодируют белки с аналогичными функциональными доменами База данных углеводно-активных ферментов (CAZy); (Lombard et al., 2014), а также определить, могут ли определенные семейства, такие как гликозилтрансферазы (GTs), гликозилгидролазы (GHs) и другие ферменты, модифицирующие углеводы, играть ключевую роль в синтезе и модификации клеточной стенки во время стресса.Поэтому после первых двух разделов этого обзора, где мы рассмотрели новые и исторические данные о роли полимеров и генов, связанных с клеточной стенкой, во время биотического и абиотического стресса, мы пересмотрели несколько наборов транскрипционных данных, чтобы обобщить реакцию клеточной стенки и гены и семейства, связанные с углеводами, при стрессе, подчеркивая замечательный уровень сохранения реакций, вызванных различными типами стресса.
Биотический стресс и клеточная стенка растений
На поле совместной эволюции между растениями и микробами на протяжении миллионов лет растения выработали многоуровневую систему защиты, в которой клеточная стенка служит множеству целей. Растительная клеточная стенка может служить предварительно сформированным или пассивным структурным барьером, а также индуцированным или активным защитным барьером. Микробы должны обойти клеточную стенку и другие заранее сформированные барьеры, чтобы установить желаемые патогенные отношения с растениями-хозяевами. Для этого требуются соответствующие стратегии распознавания хозяина и разработка подходящих инфекционных структур и/или химического оружия (Zentmyer, 1961; Turrà et al., 2015). Неспособность разработать соответствующие стратегии для разрушения стенки хозяина и других предварительно сформированных структур приводит к тому, что микробы становятся непатогенными и неадаптированными патогенами.
Растительная клеточная стенка может служить предварительно сформированным или пассивным структурным барьером, а также индуцированным или активным защитным барьером. Микробы должны обойти клеточную стенку и другие заранее сформированные барьеры, чтобы установить желаемые патогенные отношения с растениями-хозяевами. Для этого требуются соответствующие стратегии распознавания хозяина и разработка подходящих инфекционных структур и/или химического оружия (Zentmyer, 1961; Turrà et al., 2015). Неспособность разработать соответствующие стратегии для разрушения стенки хозяина и других предварительно сформированных структур приводит к тому, что микробы становятся непатогенными и неадаптированными патогенами.
Растение-хозяин также может использовать клеточную стенку в качестве активного защитного барьера для тех микробов, которые развили механизм преодоления ранее сформированных барьеров. Во время инфекции элиситоры олигосахаридов высвобождаются из клеточной стенки растения-хозяина (ассоциированные с повреждением молекулярные структуры, DAMP) или из клеточной стенки патогена (ассоциированные с патогенами молекулярные структуры, PAMP) в результате деградации (Boller and Felix, 2009). Растения воспринимают эти элиситоры через иммунные рецепторы плазматической мембраны, которые запускают сигнальные каскады для активации многочисленных защитных реакций, называемых иммунитетом, запускаемым DAMP или PAMP (DTI или PTI; Jones and Dangl, 2006).Одним из распространенных защитных ответов, связанных с DTI или PTI, является укрепление клеточной стенки для создания большей устойчивости к физическому давлению и/или ферментативному гидролизу, генерируемому патогенами (Boller and Felix, 2009; Ringli, 2010; Malinovsky et al., 2014). . В зависимости от типа взаимодействия процесс укрепления клеточной стенки может происходить несколькими различными способами, включая перестройку и сшивание ранее существовавших материалов клеточной стенки, включение легко сшиваемых полимеризованных материалов в существующую клеточную стенку и локальное отложение клеточной стенки. материалов в местах заражения (Moerschbacher and Mendgen, 2012).
Растения воспринимают эти элиситоры через иммунные рецепторы плазматической мембраны, которые запускают сигнальные каскады для активации многочисленных защитных реакций, называемых иммунитетом, запускаемым DAMP или PAMP (DTI или PTI; Jones and Dangl, 2006).Одним из распространенных защитных ответов, связанных с DTI или PTI, является укрепление клеточной стенки для создания большей устойчивости к физическому давлению и/или ферментативному гидролизу, генерируемому патогенами (Boller and Felix, 2009; Ringli, 2010; Malinovsky et al., 2014). . В зависимости от типа взаимодействия процесс укрепления клеточной стенки может происходить несколькими различными способами, включая перестройку и сшивание ранее существовавших материалов клеточной стенки, включение легко сшиваемых полимеризованных материалов в существующую клеточную стенку и локальное отложение клеточной стенки. материалов в местах заражения (Moerschbacher and Mendgen, 2012).
Состав сосочков и биотрофные патогены
Локальное отложение материалов клеточной стенки, также известное как сосочки, является ранней защитной реакцией, обычно формируемой против инфекции рядом биотрофных, гемибиотрофных и бактериальных патогенов (Bellincampi et al. , 2014). Крошечная структура микрометрового масштаба, образующаяся в месте заражения, часто бывает достаточно большой, чтобы остановить проникновение грибка. У некоторых видов-нехозяев и хозяев резистентность достигается на стадии до инвазии за счет образования сосочков в местах заражения.Однако точная роль сосочков не совсем понятна. Они могут действовать как физический барьер, который эффективно останавливает проникновение патогенов или замедляет процесс проникновения, так что другие защитные механизмы могут быть активированы раньше времени (Stone and Clarke, 1992; Huckelhoven, 2005). Они также могут функционировать в качестве химического барьера, вмещающего различные виды химического оружия, такие как противомикробные токсины, фитоалексины и дефензины, которые необходимы для непосредственной атаки патогенов или ингибирования ферментов, разрушающих клеточную стенку, вырабатываемых патогенами (Albersheim et al., 2011).
, 2014). Крошечная структура микрометрового масштаба, образующаяся в месте заражения, часто бывает достаточно большой, чтобы остановить проникновение грибка. У некоторых видов-нехозяев и хозяев резистентность достигается на стадии до инвазии за счет образования сосочков в местах заражения.Однако точная роль сосочков не совсем понятна. Они могут действовать как физический барьер, который эффективно останавливает проникновение патогенов или замедляет процесс проникновения, так что другие защитные механизмы могут быть активированы раньше времени (Stone and Clarke, 1992; Huckelhoven, 2005). Они также могут функционировать в качестве химического барьера, вмещающего различные виды химического оружия, такие как противомикробные токсины, фитоалексины и дефензины, которые необходимы для непосредственной атаки патогенов или ингибирования ферментов, разрушающих клеточную стенку, вырабатываемых патогенами (Albersheim et al., 2011).
Было высказано предположение, что резистентность к проникновению, опосредованная сосочками, представляет собой способность генотипа хозяина развивать эффективные сосочки с правильным составом и в нужное время (Aist and Israel, 1977; Inoue et al. , 1994). Следовательно, понимание состава сосочков и факторов, влияющих на развитие эффективных сосочков, было в центре внимания многих исследователей. В последние три десятилетия в исследованиях предпринимались попытки идентифицировать компоненты сосочков, образующихся против разных биотрофных патогенов на разных культурах.В то время как некоторые физико-химические изменения, которые происходят во время укрепления клеточных стенок, опосредованного сосочками, в настоящее время хорошо описаны, многие аспекты плохо изучены. Например, было проведено множество исследований, посвященных накоплению и лигнификации папиллярной каллозы из-за наличия флуоресцентных красителей для каллозы и присущей аутофлуоресценции соединений лигнина, в то время как потенциальная роль многих других компонентов клеточной стенки остается неизвестной.
, 1994). Следовательно, понимание состава сосочков и факторов, влияющих на развитие эффективных сосочков, было в центре внимания многих исследователей. В последние три десятилетия в исследованиях предпринимались попытки идентифицировать компоненты сосочков, образующихся против разных биотрофных патогенов на разных культурах.В то время как некоторые физико-химические изменения, которые происходят во время укрепления клеточных стенок, опосредованного сосочками, в настоящее время хорошо описаны, многие аспекты плохо изучены. Например, было проведено множество исследований, посвященных накоплению и лигнификации папиллярной каллозы из-за наличия флуоресцентных красителей для каллозы и присущей аутофлуоресценции соединений лигнина, в то время как потенциальная роль многих других компонентов клеточной стенки остается неизвестной.
Недавняя разработка антител, специфичных для клеточной стенки, модулей связывания углеводов и красителей с малыми молекулами дает новую возможность получить информацию о трехмерных изменениях полисахаридов в инфицированных участках клеточной стенки. Чоудхури и др. (2014) использовали эти новые инструменты, чтобы показать, что основные полисахариды, обнаруженные в сосочках ячменя, индуцируются в ответ на грибковый патоген Blumeria graminis f.sp. hordei ( Bgh ) представляют собой каллозу, арабиноксилан и целлюлозу. Эффективные сосочки, успешно предотвращающие попытки проникновения Bgh , содержат значительно более высокие концентрации этих полисахаридов по сравнению с неэффективными сосочками. Сосочки состоят из внутреннего ядра, состоящего из каллозы и арабиноксилана, и внешнего слоя, содержащего арабиноксилан и целлюлозу.Сочетание арабиноксилана и целлюлозы с устойчивостью к проникновению открывает новые возможности для улучшения состава сосочков и создания линий с улучшенной устойчивостью к болезням. Предыдущие исследования описывали профили экспрессии генов-кандидатов во время формирования сосочков и обсуждали их вероятные защитные функции (Bhuiyan et al., 2009). Однако помимо участия семейства генов , подобного глюкансинтазе ( GSL ), в синтезе папиллярной каллозы (Jacobs et al.
Чоудхури и др. (2014) использовали эти новые инструменты, чтобы показать, что основные полисахариды, обнаруженные в сосочках ячменя, индуцируются в ответ на грибковый патоген Blumeria graminis f.sp. hordei ( Bgh ) представляют собой каллозу, арабиноксилан и целлюлозу. Эффективные сосочки, успешно предотвращающие попытки проникновения Bgh , содержат значительно более высокие концентрации этих полисахаридов по сравнению с неэффективными сосочками. Сосочки состоят из внутреннего ядра, состоящего из каллозы и арабиноксилана, и внешнего слоя, содержащего арабиноксилан и целлюлозу.Сочетание арабиноксилана и целлюлозы с устойчивостью к проникновению открывает новые возможности для улучшения состава сосочков и создания линий с улучшенной устойчивостью к болезням. Предыдущие исследования описывали профили экспрессии генов-кандидатов во время формирования сосочков и обсуждали их вероятные защитные функции (Bhuiyan et al., 2009). Однако помимо участия семейства генов , подобного глюкансинтазе ( GSL ), в синтезе папиллярной каллозы (Jacobs et al. , 2003) гены, участвующие в синтезе остальных полисахаридов сосочков, еще не охарактеризованы.
, 2003) гены, участвующие в синтезе остальных полисахаридов сосочков, еще не охарактеризованы.
Некротрофические патогены
Взаимодействие растительных и некротрофных патогенов происходит на более высоком уровне, чем наблюдаемое с биотрофными патогенами. Хотя целью растительной клетки по-прежнему является предотвращение проникновения патогенов, патогены с некротрофической фазой своего жизненного цикла развили арсенал ферментов, разрушающих клеточную стенку, предназначенных для разрушения клеточной стенки растения, наряду с рядом факторов вирулентности или токсинов. чтобы убить клетки-хозяева и высвободить питательные вещества внутри, а не забрать их тайком (van Kan, 2006).Растения обычно реагируют на некротрофные патогены сильнее, но аналогично биотрофным патогенам, укрепляя клеточную стенку в месте атаки и модифицируя клеточную стенку, чтобы она была более устойчивой к ферментативному перевариванию. Патоген часто использует этот процесс в своих интересах, заставляя растение изменять свою клеточную стенку, чтобы сделать ее более усвояемой (Hok et al. , 2010). Учитывая широко распространенное повреждение, которое может быть вызвано токсинами, можно было бы также ожидать серьезного ранящего ответа из-за потери целостности клеточной стенки (Ferrari et al., 2013). Большинство некротрофных патогенов проникает в ткани растений через устьица и открытые раны, распространяясь между клеточными контактами.
, 2010). Учитывая широко распространенное повреждение, которое может быть вызвано токсинами, можно было бы также ожидать серьезного ранящего ответа из-за потери целостности клеточной стенки (Ferrari et al., 2013). Большинство некротрофных патогенов проникает в ткани растений через устьица и открытые раны, распространяясь между клеточными контактами.
Нафиси и др. (2015) рассмотрели роль клеточной стенки во взаимодействии растение:некротроф, сосредоточив внимание на передаче сигналов фитогормонов ниже по течению. Распознавание сигналов PAMP приводит к активации сигнальных каскадов, которые взаимодействуют с ауксином, цитокинином, брассиностероидами и абсцизовой кислотой, чтобы активировать экспрессию генов, связанных с защитой.Таким образом, восприимчивость клеточной стенки к деградации и последующему производству PAMP важна для устойчивости растения к патогенам. Об этом свидетельствует влияние метилэстерификации пектина на взаимодействие растений и патогенов. Лионетти и др. (2012) рассмотрели роль пектинметилэстераз в ответ на ряд патогенов растений, включая некротрофы, подчеркнув, что деэтерификация пектина влияет на восприимчивость клеточной стенки к ферментам, разрушающим клеточную стенку грибов. Был проведен метаанализ ферментов, модифицирующих пектин, у арабидопсиса, однако семейства гликозилтрансфераз, участвующие в синтезе пектина, не были включены.
Был проведен метаанализ ферментов, модифицирующих пектин, у арабидопсиса, однако семейства гликозилтрансфераз, участвующие в синтезе пектина, не были включены.
Нематоды, паразитирующие на растениях
Ремоделирование клеточной стенки при заражении паразитическими нематодами корней растений, вероятно, является важным компонентом для успешного завершения жизненного цикла нематод (обзор в Bohlmann and Sobczak, 2014). Паразитические нематоды должны проникать, мигрировать и устанавливать питательные структуры (синцитии или гигантские клетки), все из которых требуют определенного взаимодействия со стенками клеток корня.Ранние исследования цистообразующих нематод изучали роль ферментов, разрушающих клеточную стенку, которые секретируются для того, чтобы проникнуть и мигрировать к оптимальному месту питания (обзор в Deubert and Rohde, 1971), а недавние исследования подтвердили, что коктейль из ферментов, таких как целлюлазы, 1,3-β-глюканазы и пектинлиазы, обычно связанные с патогенезом растений, сохраняются у разных видов паразитических нематод (Rai et al. , 2015). Совсем недавно акцент сместился на реакцию клеточной стенки растений, поскольку она ремоделируется, чтобы приспособиться к формированию места питания (обзор в Wieczorek, 2015) и различиям, наблюдаемым в восприимчивом и резистентном взаимодействии.Несколько исследований показали специфические изменения полисахаридов стенки, таких как пектин, во время инфекции (Davies and Urwin, 2012) и выдвинули гипотезу о том, что компоненты клеточной стенки, такие как 1,3-β-глюкан или 1,3:1,4-β- глюкан может влиять на поток растворенных веществ между нематодой и хозяином (Hofmann et al., 2010; Aditya et al., 2015).
, 2015). Совсем недавно акцент сместился на реакцию клеточной стенки растений, поскольку она ремоделируется, чтобы приспособиться к формированию места питания (обзор в Wieczorek, 2015) и различиям, наблюдаемым в восприимчивом и резистентном взаимодействии.Несколько исследований показали специфические изменения полисахаридов стенки, таких как пектин, во время инфекции (Davies and Urwin, 2012) и выдвинули гипотезу о том, что компоненты клеточной стенки, такие как 1,3-β-глюкан или 1,3:1,4-β- глюкан может влиять на поток растворенных веществ между нематодой и хозяином (Hofmann et al., 2010; Aditya et al., 2015).
Травоядные насекомые
Реакция растений на нападение травоядных насекомых в значительной степени регулируется реакцией на ранение, вызванной распознаванием DAMP (Boller and Felix, 2009).Механические повреждения, вызванные кормлением насекомыми, могут быть уменьшены за счет утолщения клеточной стенки, однако устойчивость, скорее всего, примет форму химической защиты, такой как фенолы, алкалоиды, терпеноиды или глюкозинолаты (обзор в van Dam, 2009). Прямое нацеливание растительных гликозилгидролаз или лектинов на хитин или другие углеводные структуры, присутствующие в структурах питания насекомых и средней кишке, играет важную роль в защите травоядных, препятствуя поглощению патогена питательными веществами (рассмотрено в Vandenborre et al., 2011).
Прямое нацеливание растительных гликозилгидролаз или лектинов на хитин или другие углеводные структуры, присутствующие в структурах питания насекомых и средней кишке, играет важную роль в защите травоядных, препятствуя поглощению патогена питательными веществами (рассмотрено в Vandenborre et al., 2011).
Влияние измененного состава клеточной стенки на патогенез
Трансгенные и генетические подходы как с усилением, так и с потерей функции использовались для изучения влияния измененного состава стенки на устойчивость растений к болезням, некоторые из которых приведены в таблице 1. Эти исследования показывают, что модифицированный состав клеточной стенки действительно может привести к к увеличению или уменьшению фенотипов устойчивости к болезням у растений-хозяев, в зависимости от полисахарида-мишени и от того, был ли ген, связанный с клеточной стенкой, сверхэкспрессирован или мутирован.Во многих случаях гены-мишени, связанные с клеточной стенкой, были идентифицированы с помощью транскриптомных методов после применения специфического биотического стресса (см.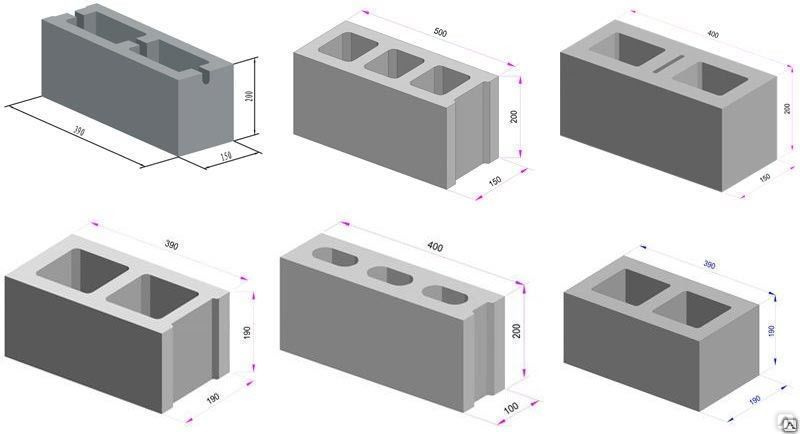 ссылки в таблицах 1, 2). Однако также важно отметить, что ряд этих исследований был направлен на улучшение усвояемости кормовых культур, чтобы сделать лигноцеллюлозу менее неподатливой для биопереработки, и есть некоторые опасения, что растения с повышенной усвояемостью из-за измененных свойств клеточных стенок могут быть более восприимчивы. к вредителям и болезням.Данные исследований трансгенных линий с измененными уровнями транскриптов генов-кандидатов, участвующих в путях биосинтеза целлюлозы, нецеллюлозных полисахаридов и лигнина, предполагают, что это может быть не так. Например, снижение биосинтеза целлюлозы с помощью генетики или химических веществ приводит к компенсаторным эффектам целостности клеточной стенки, что приводит к увеличению лигнификации и повышению устойчивости к болезням (Hamann, 2012).
ссылки в таблицах 1, 2). Однако также важно отметить, что ряд этих исследований был направлен на улучшение усвояемости кормовых культур, чтобы сделать лигноцеллюлозу менее неподатливой для биопереработки, и есть некоторые опасения, что растения с повышенной усвояемостью из-за измененных свойств клеточных стенок могут быть более восприимчивы. к вредителям и болезням.Данные исследований трансгенных линий с измененными уровнями транскриптов генов-кандидатов, участвующих в путях биосинтеза целлюлозы, нецеллюлозных полисахаридов и лигнина, предполагают, что это может быть не так. Например, снижение биосинтеза целлюлозы с помощью генетики или химических веществ приводит к компенсаторным эффектам целостности клеточной стенки, что приводит к увеличению лигнификации и повышению устойчивости к болезням (Hamann, 2012).
Таблица 1. Растение: фенотипы устойчивости к биотическому стрессу с измененным составом клеточной стенки .
Таблица 2. Растение:системы стресса, собранные из PLEXdb для метаанализа .
Абиотический стресс
Другой тип внешнего раздражителя, который может повлиять на клеточную стенку растения, — это абиотический стресс. Этот тип стресса включает ряд факторов, таких как экстремальные температуры, засуха, наводнения, соленость, атмосферные загрязнители и загрязнители тяжелыми металлами. Часто растение одновременно подвергается нескольким абиотическим стрессам, что может затруднить определение того, какой стресс вызывает наблюдаемую реакцию.Различные изменения в составе клеточных стенок растений при различных абиотических стрессах были изучены и недавно подробно рассмотрены. Ван и др. (2016) обсудили влияние четырех типов абиотического стресса; солевой стресс, доступность воды, световые условия и температура, на одном из аспектов клеточной стенки растений, целлюлозе. Среди подробно обсуждаемых генов есть члены семейства генов CesA , которые, как известно, синтезируют целлюлозу, и другие, которые ранее были идентифицированы как взаимодействующие с CesA s. Ле Галл и др. (2015) дают обзор влияния засухи, жары, холода, солей, тяжелых металлов, света и загрязнителей воздуха на основные компоненты клеточной стенки растений как у однодольных, так и у двудольных растений. Напротив, Tenhaken (2014) сосредоточился на влиянии активных форм кислорода (АФК), которые являются реакцией растений на стресс, на компоненты клеточной стенки растений, такие как XTH и экспансины. Члены семейств генов экспансина и XTH часто проявляют дифференциальную экспрессию в условиях абиотического стресса и, следовательно, повышенное присутствие АФК, что приводит к потенциальной паузе в росте.В этом разделе текущего обзора (и в таблице 3) мы приводим краткий обзор исследований, раскрывающих динамику транскрипции генов клеточной стенки во время абиотического стресса, прежде чем сосредоточить внимание на исследованиях однодольных и двудольных растений, которые напрямую связывают влияние ген или семейство генов, связанных с клеточной стенкой, к измененным реакциям на абиотический стресс.
Ле Галл и др. (2015) дают обзор влияния засухи, жары, холода, солей, тяжелых металлов, света и загрязнителей воздуха на основные компоненты клеточной стенки растений как у однодольных, так и у двудольных растений. Напротив, Tenhaken (2014) сосредоточился на влиянии активных форм кислорода (АФК), которые являются реакцией растений на стресс, на компоненты клеточной стенки растений, такие как XTH и экспансины. Члены семейств генов экспансина и XTH часто проявляют дифференциальную экспрессию в условиях абиотического стресса и, следовательно, повышенное присутствие АФК, что приводит к потенциальной паузе в росте.В этом разделе текущего обзора (и в таблице 3) мы приводим краткий обзор исследований, раскрывающих динамику транскрипции генов клеточной стенки во время абиотического стресса, прежде чем сосредоточить внимание на исследованиях однодольных и двудольных растений, которые напрямую связывают влияние ген или семейство генов, связанных с клеточной стенкой, к измененным реакциям на абиотический стресс.
Таблица 3. Растение: фенотип реакции на абиотический стресс с измененным составом клеточной стенки .
Глобальное профилирование реакции на абиотический стресс
Транскрипционные изменения, сопровождающие различные абиотические стрессы, обсуждались довольно подробно (обзор в Santos et al., 2011; Gehan et al., 2015), но очень немногие рассматривали эти изменения в контексте специфических генов, связанных с клеточной стенкой. Детальный анализ генетических реакций на засуху отдельных органов колоса ячменя был проведен Abebe et al. (2010). Профили транскрипции ости, семени, цветковой чешуи и чешуи сравнивали между растениями, которые подвергались засушливому стрессу из-за отсутствия воды в течение 4 дней во время налива зерна, и контрольными растениями. Было обнаружено, что для всех тканей, кроме семян, множественные гены, связанные с клеточной стенкой, по-разному регулируются между контрольными растениями и растениями, подвергшимися засухе. Гены, кодирующие членов семейств целлюлозосинтазы (GT2, CesA), UDP-ксилозилтрансферазы, семейства гликозилгидролаз 1 (Gh2), эндо-бета-1,4-глюканазы (GH9) и ксилоглюканэндотрансгликозилазы (Gh26, XTH/XET), были среди генов, связанных с клеточной стенкой, наблюдается подавление в условиях засухи. Дополнительный XET активировался в условиях засухи, а также предполагаемый ингибитор ксиланазы, эндо-1,3-бета-глюкозидаза и бета-D-глюканэкзогидролаза. Аналогичные исследования у арабидопсиса выявили более 500 генов, которые реагируют на засуху, холод и засоленность (Seki et al., 2002), в том числе несколько представителей семейств экстенсинов, пектинэстераз и XTH/XET, которые были подавлены. Ван и др. (2013) показали, что только в случае засоления более 140 генов, связанных с клеточной стенкой, реагируют на солевой стресс, а иногда и по-разному между экотипами Arabidopsis. Как ранее было установлено другими авторами в условиях засухи (Wu and Cosgrove, 2000; Moore et al., 2008), в зависимости от того, какая ткань наблюдается, клеточная стенка растения либо ослабевает, либо сжимается для поддержания роста.
Гены, кодирующие членов семейств целлюлозосинтазы (GT2, CesA), UDP-ксилозилтрансферазы, семейства гликозилгидролаз 1 (Gh2), эндо-бета-1,4-глюканазы (GH9) и ксилоглюканэндотрансгликозилазы (Gh26, XTH/XET), были среди генов, связанных с клеточной стенкой, наблюдается подавление в условиях засухи. Дополнительный XET активировался в условиях засухи, а также предполагаемый ингибитор ксиланазы, эндо-1,3-бета-глюкозидаза и бета-D-глюканэкзогидролаза. Аналогичные исследования у арабидопсиса выявили более 500 генов, которые реагируют на засуху, холод и засоленность (Seki et al., 2002), в том числе несколько представителей семейств экстенсинов, пектинэстераз и XTH/XET, которые были подавлены. Ван и др. (2013) показали, что только в случае засоления более 140 генов, связанных с клеточной стенкой, реагируют на солевой стресс, а иногда и по-разному между экотипами Arabidopsis. Как ранее было установлено другими авторами в условиях засухи (Wu and Cosgrove, 2000; Moore et al., 2008), в зависимости от того, какая ткань наблюдается, клеточная стенка растения либо ослабевает, либо сжимается для поддержания роста. Это иллюстрирует сложность реакции клеточной стенки на абиотические стрессы.
Это иллюстрирует сложность реакции клеточной стенки на абиотические стрессы.
Растения могут испытывать абиотический стресс разной степени тяжести, поэтому многие исследования включают несколько уровней лечения стресса, чтобы понять, как это влияет на реакцию. Харб и др. (2010) оценили влияние прогрессирующей засухи и умеренной засухи на рост растений с помощью ряда биохимических и физиологических анализов, а изменения в экспрессии генов отслеживали с помощью эксперимента с микроматрицами.В условиях умеренной засухи рост растений значительно снижался как по накоплению биомассы, так и по распусканию листьев, а также по устьичной проводимости. В этих условиях несколько генов, кодирующих экспансины клеточной стенки, активировались; однако в условиях прогрессирующей засухи экспансины клеточной стенки подавлялись. Экспансины представляют собой белки, которые, как было показано ранее, ослабляют и модифицируют клеточную стенку растений во время роста и адаптации к стрессу путем модификации целлюлозных и нецеллюлозных компонентов клеточной стенки (Cosgrove, 2005). Применяя аналогичный подход, Mangelsen et al. (2011) подвергали молодые зерновки ячменя тепловому стрессу в течение 0,5, 3 и 6 часов и использовали микрочипы для идентификации дифференциально экспрессируемых генов. Гены с пониженной регуляцией, связанные с клеточной стенкой, были статистически чрезмерно представлены, особенно после 3 и 6 часов воздействия теплового стресса, которые авторы описали как фазы первичной тепловой реакции и адаптации к тепловому стрессу соответственно. Этот набор включал гены, функционально аннотированные как пектатлиазы, полигалактуроназы и пектинэстеразы.
Применяя аналогичный подход, Mangelsen et al. (2011) подвергали молодые зерновки ячменя тепловому стрессу в течение 0,5, 3 и 6 часов и использовали микрочипы для идентификации дифференциально экспрессируемых генов. Гены с пониженной регуляцией, связанные с клеточной стенкой, были статистически чрезмерно представлены, особенно после 3 и 6 часов воздействия теплового стресса, которые авторы описали как фазы первичной тепловой реакции и адаптации к тепловому стрессу соответственно. Этот набор включал гены, функционально аннотированные как пектатлиазы, полигалактуроназы и пектинэстеразы.
В других подходах сравнивали транскриптомные данные чувствительных к стрессу и толерантных сортов. Кэл и др. (2013) сравнили данные транскриптома для зоны удлинения листа (LEZ) двух сортов риса, Moroberekan, который является засухоустойчивым, и IR64, который чувствителен к засухе, в условиях водного дефицита. Эта ткань была выбрана потому, что изменения в экспансии в LEZ часто являются одним из более ранних ответов на дефицит воды (Cutler et al. , 1980). Эти наборы транскриптомных данных идентифицировали набор генов, которые показали более чем 2-кратное изменение экспрессии в обоих сортах, включая 27 генов, связанных с клеточной стенкой, большинство из которых были подавлены в засухоустойчивом сорте Мороберекан.Список подавленных генов включает гены, участвующие в производстве вторичной клеточной стенки, включая циннамоил-КоА-редуктазу, ферулат-5-гидроксилазу, лакказу и апопластные пероксидазы класса III. Гены, кодирующие арабиногалактановые белки и экстенсины, участвующие в передаче сигналов и структуре клеточной стенки, XTH/XET и GT, включая два гена CesA , по-разному экспрессировались между двумя сортами. Два гена, экспрессия которых была обнаружена в Мороберекане, являются членами семейства гликозилгидролаз Gh38, кодирующего полигалактуроназы.В аналогичном исследовании Zheng et al. (2010) сравнили данные по полногеномной экспрессии генов для линий кукурузы Han21 и Ye478, устойчивых к засухе и неустойчивых к засухе, соответственно, в условиях засушливого стресса.
, 1980). Эти наборы транскриптомных данных идентифицировали набор генов, которые показали более чем 2-кратное изменение экспрессии в обоих сортах, включая 27 генов, связанных с клеточной стенкой, большинство из которых были подавлены в засухоустойчивом сорте Мороберекан.Список подавленных генов включает гены, участвующие в производстве вторичной клеточной стенки, включая циннамоил-КоА-редуктазу, ферулат-5-гидроксилазу, лакказу и апопластные пероксидазы класса III. Гены, кодирующие арабиногалактановые белки и экстенсины, участвующие в передаче сигналов и структуре клеточной стенки, XTH/XET и GT, включая два гена CesA , по-разному экспрессировались между двумя сортами. Два гена, экспрессия которых была обнаружена в Мороберекане, являются членами семейства гликозилгидролаз Gh38, кодирующего полигалактуроназы.В аналогичном исследовании Zheng et al. (2010) сравнили данные по полногеномной экспрессии генов для линий кукурузы Han21 и Ye478, устойчивых к засухе и неустойчивых к засухе, соответственно, в условиях засушливого стресса. В общей сложности 15 наборов зондов, которые кодируют предполагаемые гены, связанные с клеточной стенкой, по-разному экспрессировались между двумя линиями. К ним относятся зонды, аннотированные как субъединицы целлюлозосинтазы, предшественники эндо-1,3-β-глюкозидазы и предшественники белка COBRA-подобного 3.
В общей сложности 15 наборов зондов, которые кодируют предполагаемые гены, связанные с клеточной стенкой, по-разному экспрессировались между двумя линиями. К ним относятся зонды, аннотированные как субъединицы целлюлозосинтазы, предшественники эндо-1,3-β-глюкозидазы и предшественники белка COBRA-подобного 3.
Как упоминалось ранее в этом обзоре, растения часто одновременно подвергаются множественным стрессам, включая как абиотические, так и биотические, и их эффекты не обязательно являются просто аддитивными (Puranik et al., 2012; Кулен и др., 2016). Поэтому важно, несмотря на очевидную сложность подобных экспериментов, изучить влияние на растения многократных одновременных стрессов. Наблюдалась активация AtRALFL8 в корнях 10-дневных растений при применении двойного стресса инфекции нематодами и водного дефицита (Atkinson et al., 2013). Последующий микрочиповый анализ показал, что в этих условиях AtRALFL8 в высокой степени коэкспрессируется с пектиназами, известными своей способностью способствовать ремоделированию клеточной стенки, ранее было показано, что они играют роль в нескольких ответных реакциях на стресс, включая заражение нематодами (Pelloux et al. , 2007; Ан и др., 2008). Кулен и др. (2016) подвергали растения арабидопсиса одиночному и двойному сочетанию стресса от засухи, грибковой инфекции Botrytis cinerea и травоядной инфекции Pieris rapae . В общей сложности 41 ген, связанный с клеточной стенкой, включая PECTIN METHYLESTERASE 3 ( PME3 ), EXPANSIN A6 ( EXPA6 ), XTh20 и XTh42 , отвечает Целлюлоза синтазы, такая как G2 ( CSLG2 ), Arabinogalactan белок 2 ( AGP2 ), Beta Glucosidase 46 ( BGLU46 ), 1,3-β-глюканаза и Excansin A8 ( EXPA8 ) ответил одинаково на все три.Это указывает на то, что общие транскрипционные ответы и, возможно, последующие эффекты на состав клеточной стенки используются в ответ на разные стрессы.
, 2007; Ан и др., 2008). Кулен и др. (2016) подвергали растения арабидопсиса одиночному и двойному сочетанию стресса от засухи, грибковой инфекции Botrytis cinerea и травоядной инфекции Pieris rapae . В общей сложности 41 ген, связанный с клеточной стенкой, включая PECTIN METHYLESTERASE 3 ( PME3 ), EXPANSIN A6 ( EXPA6 ), XTh20 и XTh42 , отвечает Целлюлоза синтазы, такая как G2 ( CSLG2 ), Arabinogalactan белок 2 ( AGP2 ), Beta Glucosidase 46 ( BGLU46 ), 1,3-β-глюканаза и Excansin A8 ( EXPA8 ) ответил одинаково на все три.Это указывает на то, что общие транскрипционные ответы и, возможно, последующие эффекты на состав клеточной стенки используются в ответ на разные стрессы.
Генетические и трансгенные данные, подтверждающие роль генов клеточной стенки в реакции на абиотический стресс
Подобно тому, что описано для биотических стрессов, мутантные линии были ценным ресурсом с точки зрения понимания того, как гены, связанные с клеточной стенкой, могут смягчать или усиливать реакцию на абиотический стресс. У арабидопсиса AtCesA8 / IRX1 способствует синтезу вторичной клеточной стенки и влияет на устойчивость растений к засухе и осмотическому стрессу (Chen et al., 2005). Мутантные аллели AtCesA8, увядание листьев 2-1 ( lew2-1 ) и lew2-2 показали более высокую устойчивость к осмотическому стрессу, индуцированному воздействием NaCl и маннита, и стрессу засухи по сравнению с растениями дикого типа. . Другие компоненты клеточной стенки растений влияют на устойчивость растений к холоду или морозу. Например, Такета и др. (2012) провели скрининг мутантов ячменя, индуцированных азидом натрия, которые были чувствительны к охлаждению. Было обнаружено, что из 11 идентифицированных линий 2 лишены (1,3:1,4)-β-глюкана и содержат мутации HvCslF6 , связанного с клеточной стенкой гена, который ранее был вовлечен в синтез (1, 3:1,4)-β-глюкан (Burton et al., 2006). Хотя эти линии не содержали мутаций, которые продуцировали бы преждевременный стоп-кодон, одна мутация располагалась в непосредственной близости от консервативного каталитического мотива HvCslF6 гликозилтрансферазы (GT2).
У арабидопсиса AtCesA8 / IRX1 способствует синтезу вторичной клеточной стенки и влияет на устойчивость растений к засухе и осмотическому стрессу (Chen et al., 2005). Мутантные аллели AtCesA8, увядание листьев 2-1 ( lew2-1 ) и lew2-2 показали более высокую устойчивость к осмотическому стрессу, индуцированному воздействием NaCl и маннита, и стрессу засухи по сравнению с растениями дикого типа. . Другие компоненты клеточной стенки растений влияют на устойчивость растений к холоду или морозу. Например, Такета и др. (2012) провели скрининг мутантов ячменя, индуцированных азидом натрия, которые были чувствительны к охлаждению. Было обнаружено, что из 11 идентифицированных линий 2 лишены (1,3:1,4)-β-глюкана и содержат мутации HvCslF6 , связанного с клеточной стенкой гена, который ранее был вовлечен в синтез (1, 3:1,4)-β-глюкан (Burton et al., 2006). Хотя эти линии не содержали мутаций, которые продуцировали бы преждевременный стоп-кодон, одна мутация располагалась в непосредственной близости от консервативного каталитического мотива HvCslF6 гликозилтрансферазы (GT2). Такета и др. (2012) предположили, что повышенная чувствительность вегетативных тканей к охлаждению у линий, содержащих мутации HvCslF6 , может быть связана с более тонкими клеточными стенками, поскольку (1,3:1,4)-β-глюкан обычно является основным компонентом этого структура в травах. Эта гипотеза была подкреплена работой Cu et al.(2016), которые наблюдали более тонкие клеточные стенки в линиях нокдауна CslF6 , генерируемых РНКи, по сравнению с диким типом, используя как метод окрашивания Calcofluor, так и иммуноцитологическое окрашивание с помощью BG1 (1,3:1,4)-β-глюкан-специфического антитела. . Интересно, что содержание (1,3:1,4)-β-глюкана в зерне злаков, по-видимому, особенно чувствительно к условиям окружающей среды, хотя неясно, зависит ли это изменение только от модифицированной функции HvCslF6 . Суонстон и др. (1997) и Wallwork et al.(1998) выявили значительные различия в содержании (1,3:1,4)-β-глюкана в зерне ячменя в зависимости от участка поля или температуры во время созревания зерна.
Такета и др. (2012) предположили, что повышенная чувствительность вегетативных тканей к охлаждению у линий, содержащих мутации HvCslF6 , может быть связана с более тонкими клеточными стенками, поскольку (1,3:1,4)-β-глюкан обычно является основным компонентом этого структура в травах. Эта гипотеза была подкреплена работой Cu et al.(2016), которые наблюдали более тонкие клеточные стенки в линиях нокдауна CslF6 , генерируемых РНКи, по сравнению с диким типом, используя как метод окрашивания Calcofluor, так и иммуноцитологическое окрашивание с помощью BG1 (1,3:1,4)-β-глюкан-специфического антитела. . Интересно, что содержание (1,3:1,4)-β-глюкана в зерне злаков, по-видимому, особенно чувствительно к условиям окружающей среды, хотя неясно, зависит ли это изменение только от модифицированной функции HvCslF6 . Суонстон и др. (1997) и Wallwork et al.(1998) выявили значительные различия в содержании (1,3:1,4)-β-глюкана в зерне ячменя в зависимости от участка поля или температуры во время созревания зерна. Аналогичным образом, у нескольких сортов пшеницы, выращенных в различных условиях жары и засухи, наблюдалось снижение содержания (1,3:1,4)-β-глюкана в зерне у линий, выращенных в стрессовых условиях (Rakszegi et al., 2014). И наоборот, в тех же условиях сообщалось об увеличении содержания арабиноксилана в зерне, что, возможно, способствовало снижению содержания (1,3:1,4)-β-глюкана.Генетическая основа этих переменных реакций на абиотический стресс еще не раскрыта.
Аналогичным образом, у нескольких сортов пшеницы, выращенных в различных условиях жары и засухи, наблюдалось снижение содержания (1,3:1,4)-β-глюкана в зерне у линий, выращенных в стрессовых условиях (Rakszegi et al., 2014). И наоборот, в тех же условиях сообщалось об увеличении содержания арабиноксилана в зерне, что, возможно, способствовало снижению содержания (1,3:1,4)-β-глюкана.Генетическая основа этих переменных реакций на абиотический стресс еще не раскрыта.
Трансгенные растения также использовались для проверки роли генов-кандидатов, потенциально участвующих в производстве/модификации клеточной стенки и абиотическом стрессе. Обычно считается, что ферменты XET/XTH играют роль в ослаблении клеточной стенки и, следовательно, в расширении клеток (Rose et al., 2002). Трансгенные линии арабидопсиса, экспрессирующие XTH из Capsicum annuum , демонстрируют аномальные фенотипы листьев, включая неправильный рисунок клеток на поперечных срезах и скрученные листья (Cho et al. , 2006). Кроме того, трансгенные линии арабидопсиса и томата (Choi et al., 2011), экспрессирующие Capsicum XTH , показали повышенную солеустойчивость и более длинные корни, чем у контрольных растений, лишенных трансгена, что указывает на роль гибкости стенок в смягчении стрессовых реакций. Было обнаружено, что в тканях корней кукурузы множественные гены, связанные с клеточной стенкой, по-разному экспрессируются при солевом стрессе (Li et al., 2014), включая ZmXET1. Считается, что ZmXET1 участвует в расширении клеточной стенки, поскольку он способен гидролизовать и повторно соединять молекулы ксилоглюкана (Fry et al., 1992). Другие гены, идентифицированные Li et al. (2014) как экспансины ZmEXPA1, ZmEXPA3, ZmEXPA5, ZmEXPB1, ZmEXPB2 , активизировавшиеся, когда растения подвергались воздействию повышенной засоленности, и, следовательно, возможно участвующие в опосредовании устойчивости к токсичности, связанной с засолением. Экспрессия этих генов, связанных с клеточной стенкой, может находиться под эпигенетическим контролем, поскольку повышенная экспрессия генов ZmHATB и ZmGCN5 гистоновых ацетилтрансфераз увеличивалась после солевого стресса и сопровождалась повышенным ацетилированием гистонов h4K9 и h5K5.
, 2006). Кроме того, трансгенные линии арабидопсиса и томата (Choi et al., 2011), экспрессирующие Capsicum XTH , показали повышенную солеустойчивость и более длинные корни, чем у контрольных растений, лишенных трансгена, что указывает на роль гибкости стенок в смягчении стрессовых реакций. Было обнаружено, что в тканях корней кукурузы множественные гены, связанные с клеточной стенкой, по-разному экспрессируются при солевом стрессе (Li et al., 2014), включая ZmXET1. Считается, что ZmXET1 участвует в расширении клеточной стенки, поскольку он способен гидролизовать и повторно соединять молекулы ксилоглюкана (Fry et al., 1992). Другие гены, идентифицированные Li et al. (2014) как экспансины ZmEXPA1, ZmEXPA3, ZmEXPA5, ZmEXPB1, ZmEXPB2 , активизировавшиеся, когда растения подвергались воздействию повышенной засоленности, и, следовательно, возможно участвующие в опосредовании устойчивости к токсичности, связанной с засолением. Экспрессия этих генов, связанных с клеточной стенкой, может находиться под эпигенетическим контролем, поскольку повышенная экспрессия генов ZmHATB и ZmGCN5 гистоновых ацетилтрансфераз увеличивалась после солевого стресса и сопровождалась повышенным ацетилированием гистонов h4K9 и h5K5. В отдельном исследовании Liu et al. (2014) было обнаружено, что сверхэкспрессия OsBURP16 увеличивает количество полигалактуроназы (PG), фермента, гидролизующего пектин, и изменяет состав клеточной стенки растений. Следовательно, растения риса, сверхэкспрессирующие OsBURP16 , показали меньшую устойчивость к засухе (количественно оцениваемую как выживаемость после лишения 2-недельных растений воды), при этом растения дикого типа показали выживаемость 42% по сравнению с <10% для линий сверхэкспрессии OsBURP16 .Измерение уровней H 2 O 2 , индикатора стресса, показало, что линии OsBURP16 со сверхэкспрессией также были более восприимчивы к солевому стрессу, чем растения дикого типа.
В отдельном исследовании Liu et al. (2014) было обнаружено, что сверхэкспрессия OsBURP16 увеличивает количество полигалактуроназы (PG), фермента, гидролизующего пектин, и изменяет состав клеточной стенки растений. Следовательно, растения риса, сверхэкспрессирующие OsBURP16 , показали меньшую устойчивость к засухе (количественно оцениваемую как выживаемость после лишения 2-недельных растений воды), при этом растения дикого типа показали выживаемость 42% по сравнению с <10% для линий сверхэкспрессии OsBURP16 .Измерение уровней H 2 O 2 , индикатора стресса, показало, что линии OsBURP16 со сверхэкспрессией также были более восприимчивы к солевому стрессу, чем растения дикого типа.
Выявление сети реагирования на стресс клеточной стенки
По мере того, как мы узнаем больше о сетях генов, регулирующих синтез и гидролиз клеточной стенки растений, возможно, что путем ассоциации будет выявлено больше генов, участвующих в реакции на стресс. Недавнее подробное исследование, в котором использовались методы in vitro и in vivo для всесторонней характеристики сети генов, регулирующих синтез вторичной клеточной стенки у арабидопсиса, также показало, как на одну часть этой сети влиял абиотический стресс (Taylor-Teeples et al. др., 2015). Авторы описали регуляторную сеть ксилемы и то, как изменения солености и железа могут вызывать возмущения, которые, в свою очередь, вызывают фенотипические изменения во вторичной клеточной стенке.
Недавнее подробное исследование, в котором использовались методы in vitro и in vivo для всесторонней характеристики сети генов, регулирующих синтез вторичной клеточной стенки у арабидопсиса, также показало, как на одну часть этой сети влиял абиотический стресс (Taylor-Teeples et al. др., 2015). Авторы описали регуляторную сеть ксилемы и то, как изменения солености и железа могут вызывать возмущения, которые, в свою очередь, вызывают фенотипические изменения во вторичной клеточной стенке.
Имеется большой объем данных из предыдущих исследований абиотического стресса, которые подробно описывают глобальный ответ транскрипции генов, а в некоторых случаях — оценку изменений фенотипов клеточных стенок в качестве ответа. Аналогичные наборы данных доступны для биотических стрессов, применяемых к различным видам и тканям.Из генетических исследований, рассмотренных выше, видно, что сходные члены семейства генов (например, семейства XET/XTH, экспансинов и пектиновых модификаторов) часто участвуют в реакции на разные стрессы. Однако присущая стенке сложность и большое количество генов, участвующих в ее синтезе и модификации, означает, что многие детали в отношении генетической и биохимической основы ответа клеточной стенки на стресс остаются неясными. Несмотря на очевидные трудности, связанные со сравнением экспериментов между разными видами, стрессами и тканями, в заключительном разделе этого обзора мы пересмотрели общедоступные наборы данных транскриптома, чтобы подчеркнуть широкое сходство между различными типами стресса и подумать, можно ли уделить больше внимания предполагаемой клеточной стенке. родственные гены, которые ранее не учитывались.
Однако присущая стенке сложность и большое количество генов, участвующих в ее синтезе и модификации, означает, что многие детали в отношении генетической и биохимической основы ответа клеточной стенки на стресс остаются неясными. Несмотря на очевидные трудности, связанные со сравнением экспериментов между разными видами, стрессами и тканями, в заключительном разделе этого обзора мы пересмотрели общедоступные наборы данных транскриптома, чтобы подчеркнуть широкое сходство между различными типами стресса и подумать, можно ли уделить больше внимания предполагаемой клеточной стенке. родственные гены, которые ранее не учитывались.
Общедоступные наборы данных выделяют сложные ответы на уровне транскриптома на абиотический и биотический стресс
В предыдущих разделах этого обзора были обобщены исследования, проведенные по различным аспектам укрепления и модификации клеточной стенки во время патогенной инфекции и абиотического стресса. Укрепление клеточной стенки в виде сосочков — относительно распространенный механизм, определяющий исход инфекции. Однако, учитывая разнообразие биотических стрессов, которым может подвергаться растение, любые общие черты в формировании сосочков, вероятно, будут сопровождаться рядом различных защитных реакций, связанных с клеточной стенкой.То же самое можно ожидать и в отношении различных абиотических стрессов, таких как экстремальные температуры, соленость и наводнения. Что касается перекрытия биотических и абиотических стрессов, недавнее исследование на арабидопсисе показало, что ~ 25% транскриптов, связанных с клеточной стенкой, которые реагировали на грибковую инфекцию, растительноядность или засуху, демонстрировали сходную реакцию при каждом воздействии (Coolen et al. , 2016). Хотя в настоящее время невозможно провести подробный обзор всех изменений клеточных стенок, вызванных реакцией на ряд различных биотических и абиотических стрессов, можно провести метаанализ с использованием общедоступных данных об экспрессии транскриптов растений-патогенов и взаимодействия растений и стресса, чтобы выявить перекрытия в реакциях механизмов клеточной стенки.
Однако, учитывая разнообразие биотических стрессов, которым может подвергаться растение, любые общие черты в формировании сосочков, вероятно, будут сопровождаться рядом различных защитных реакций, связанных с клеточной стенкой.То же самое можно ожидать и в отношении различных абиотических стрессов, таких как экстремальные температуры, соленость и наводнения. Что касается перекрытия биотических и абиотических стрессов, недавнее исследование на арабидопсисе показало, что ~ 25% транскриптов, связанных с клеточной стенкой, которые реагировали на грибковую инфекцию, растительноядность или засуху, демонстрировали сходную реакцию при каждом воздействии (Coolen et al. , 2016). Хотя в настоящее время невозможно провести подробный обзор всех изменений клеточных стенок, вызванных реакцией на ряд различных биотических и абиотических стрессов, можно провести метаанализ с использованием общедоступных данных об экспрессии транскриптов растений-патогенов и взаимодействия растений и стресса, чтобы выявить перекрытия в реакциях механизмов клеточной стенки.
доступны в Базе данных экспрессии растений (PLEXdb; Dash et al., 2012), включая множество наборов данных микрочипов арабидопсиса и ячменя, в которых подробно описаны изменения в количестве транскриптов после воздействия различных абиотических или биотических стрессов (таблица 2). Дополнительным источником значительного использования является база данных Carbohydrate-Active EnZYmes (CAZy) (Lombard et al., 2014), в которой описаны семейства структурно родственных ферментов, которые гидролизуют, модифицируют или создают гликозидные связи.Используя эту информацию, были выбраны предполагаемые генов CAZy Arabidopsis , присутствующих в массиве генома Affymetrix 22K ATh2. Домены семейства белков (Pfam), связанные с аннотациями базы данных CAZy, использовались для идентификации генов, связанных с углеводами ячменя, присутствующих на 22K Barley1 GeneChip. Нормализованные уровни транскриптов для каждого гена, связанного с углеводами, из Arabidopsis и ячменя сравнивали после каждого стресса (относительно необработанных контролей) в каждом эксперименте и представляли как логарифмическую (2)-кратную индукцию. Как и следовало ожидать из предыдущих разделов этого обзора, многие гены клеточной стенки продемонстрировали выраженную реакцию на различные стрессы.
Как и следовало ожидать из предыдущих разделов этого обзора, многие гены клеточной стенки продемонстрировали выраженную реакцию на различные стрессы.
Чтобы проверить, могут ли эти ответы быть более консервативными на уровне семейства генов CAZy, была рассчитана и проанализирована средняя кратность индукции, наблюдаемая для всех членов семейства, с использованием средства просмотра мультиэкспериментов TIGR (МэВ). Иерархическая кластеризация использовалась для упорядочения семейств генов в соответствии со сходством паттерна экспрессии генов (рис. 1А, В) (Eisen et al., 1998). На рисунках 1A, B ясно показано, что большинство семейств генов CAZy активируются в ответ на абиотический или биотический стресс у Arabidopsis и ячменя. Хотя не все семейства CAZy содержат членов, которые действуют на один и тот же субстрат, и вероятность того, что все специализированные члены семейства реагируют одинаковым образом, невелика, этот подход был нацелен на предоставление простых средств идентификации ключевых видов активности, связанных с углеводами, которые являются общими для разных групп. разные стрессы. Сходное поведение хорошо охарактеризованных и плохо охарактеризованных семейств CAZy может дать полезную информацию о новых связанных со стрессом изменениях клеточной стенки и связанных с углеводами.Чтобы определить тенденции, сохраняющиеся в ответ на стрессы между Arabidopsis и ячменем, кратность индукции для каждого семейства генов была усреднена для всех абиотических и всех биотических стрессов и представлена на рисунке 1C. Сравнительные реакции этих семейств генов на абиотический и биотический стресс показаны на рисунке 2 у обоих видов.
разные стрессы. Сходное поведение хорошо охарактеризованных и плохо охарактеризованных семейств CAZy может дать полезную информацию о новых связанных со стрессом изменениях клеточной стенки и связанных с углеводами.Чтобы определить тенденции, сохраняющиеся в ответ на стрессы между Arabidopsis и ячменем, кратность индукции для каждого семейства генов была усреднена для всех абиотических и всех биотических стрессов и представлена на рисунке 1C. Сравнительные реакции этих семейств генов на абиотический и биотический стресс показаны на рисунке 2 у обоих видов.
Рисунок 1. Анализ транскриптов, связанных с клеточной стенкой, после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) .Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1. Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. Тенденции, сохраняющиеся в ответ на стрессы между арабидопсисом и ячменем, наблюдаются в (C) , который показывает среднюю кратность индукции для каждого семейства генов для всех абиотических и всех биотических стрессов у арабидопсиса и ячменя.Звездочками отмечены семейства генов, экспрессия которых усиливается как абиотическими, так и биотическими стрессами у арабидопсиса и ячменя.
Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. Тенденции, сохраняющиеся в ответ на стрессы между арабидопсисом и ячменем, наблюдаются в (C) , который показывает среднюю кратность индукции для каждого семейства генов для всех абиотических и всех биотических стрессов у арабидопсиса и ячменя.Звездочками отмечены семейства генов, экспрессия которых усиливается как абиотическими, так и биотическими стрессами у арабидопсиса и ячменя.
Рисунок 2. Графическое представление средней логарифмической (2)-кратной индукции для каждого семейства генов (представлено на рисунке 1C), которая показывает среднюю абиотическую (ось x ) и среднюю биотическую (ось y ) реакцию на стресс в Арабидопсис (А) и ячмень (В) . Семейства CAZy, которые активируются в ответ на абиотические, но не биотические стрессы, окрашены в красный цвет, семейства CAZy, которые активируются в ответ на биотические стрессы, но не в ответ на абиотические стрессы, окрашены в желтый цвет, а семейства CAZy, которые активируются в ответ как на абиотические, так и на абиотические стрессы биотические стрессы окрашены в оранжевый цвет.
Были рассмотрены два различных метода кластеризации. Внутри каждого вида семейства генов могут быть сгруппированы на основе корреляции их профилей транскриптов в каждом эксперименте. Во-вторых, наборы экспериментальных данных также могут быть сгруппированы на основе корреляции профилей транскриптов семейства генов. Интересно отметить, что абиотические стрессы обычно образуют кластер вместе, как и биотические стрессы, даже несмотря на то, что в каждом кластере существует огромная разница в типе стресса (рис. 1А, В).Догму о том, что стандартная защитная реакция клеточной стенки в первую очередь обусловлена каллозой и глюкансинтазоподобными генами семейства GT48 CAZy, трудно поддержать, учитывая большое количество семейств генов, которые, по-видимому, активируются при большинстве стрессов. Несмотря на то, что отдельные гены, индуцируемые в каждом эксперименте, различаются, кластеризация семейств CAZy в экспериментах предполагает, что существует сходная защитная реакция независимо от конкретного типа стресса. Мы можем видеть примеры биотрофных грибов, вызывающих аналогичные реакции на некротрофные грибы, стресс, вызванный засухой, вызывающий аналогичные реакции на холодовой стресс, и даже примеры в разных тканях с нематодами в корнях по сравнению с листьями, зараженными белокрылкой.
Мы можем видеть примеры биотрофных грибов, вызывающих аналогичные реакции на некротрофные грибы, стресс, вызванный засухой, вызывающий аналогичные реакции на холодовой стресс, и даже примеры в разных тканях с нематодами в корнях по сравнению с листьями, зараженными белокрылкой.
Основанная на знаниях идентификация семейств углеводов, которые реагируют на биотический и абиотический стресс
Кластеризация ответа семейств CAZy идентифицирует активности, которые, по-видимому, в большинстве случаев активируются в большинстве экспериментов, и, следовательно, клеточная стенка или углеводные компоненты могут изменяться аналогичным образом во время взаимодействия.На фигуре 1C показана средняя кратность индукции для каждого семейства генов CAZy при всех абиотических стрессах арабидопсиса, биотических стрессах арабидопсиса, абиотических стрессах ячменя и биотических стрессах ячменя. Существуют примеры семейств CAZy, которые, по-видимому, активируются (в среднем) только у Arabidopsis, включая арабиногалактановые белки (AGP), экспансины, фасциклин-подобные арабиногалактановые белки (FLA), пектинлиазу (PL1), пектин-ацетилэстеразу (CE13). , гликозилгидролазы (GH9, GH85) и ряд гликозилтрансфераз (GT4, GT20, GT21, GT47 и GT64).Было обнаружено, что число семейств CAZy, в среднем выше в ячмене, включает ксилан-ацетилэстеразу (CE6) и гликозилтрансферазу (GT14). Учитывая различия между клеточными стенками ячменя и арабидопсиса: стенки ячменя содержат больше арабиноксилана и (1,3:1,4)-β-глюкана, а стенки арабидопсиса содержат больше пектина и ксилоглюкана (Burton et al., 2010), неожиданным было увидеть большую представленность ферментов, модифицирующих пектин, в наборе данных Arabidopsis.
, гликозилгидролазы (GH9, GH85) и ряд гликозилтрансфераз (GT4, GT20, GT21, GT47 и GT64).Было обнаружено, что число семейств CAZy, в среднем выше в ячмене, включает ксилан-ацетилэстеразу (CE6) и гликозилтрансферазу (GT14). Учитывая различия между клеточными стенками ячменя и арабидопсиса: стенки ячменя содержат больше арабиноксилана и (1,3:1,4)-β-глюкана, а стенки арабидопсиса содержат больше пектина и ксилоглюкана (Burton et al., 2010), неожиданным было увидеть большую представленность ферментов, модифицирующих пектин, в наборе данных Arabidopsis.
Двенадцать семейств генов CAZy активируются (в среднем) при абиотических и биотических стрессах у арабидопсиса и ячменя.К ним относятся ферменты, расщепляющие и модифицирующие полисахариды, такие как пектинметилэстераза (CE8), углеводная эстераза (CE10) и гликозилгидролазы (Gh2, Gh27, Gh28 и Gh29), которые нацелены на ряд полисахаридов и олигосахаридов, содержащих 1,3-β-глюкан. и хитин. Некоторые из этих семейств CAZy уже вовлечены в реакцию на стресс как белки, связанные с патогенезом (PR). Гены Gh27 были классифицированы как белки PR-2, разлагающие 1,3-β-глюкан (Leubner-Metzger and Meins, 1999), в то время как Gh28 и Gh29 представляют пять из 17 семейств белков PR растений (Minic, 2008).Члены семейства Gh2 вовлечены в активацию защитных соединений посредством удаления β-глюкозида (Poulton, 1990; Duroux et al., 1998). Метилэстеразы пектина модифицируют статус этерификации пектина в стенке, влияя на восприимчивость барьера клеточной стенки к грибковым и бактериальным CWDE (Collmer and Keen, 1986). Деэстерификация пектина также влияет на пористость плазмодесм, что может изменить распространение сигнальных молекул во время защитной реакции (Chen et al., 2000).
Гены Gh27 были классифицированы как белки PR-2, разлагающие 1,3-β-глюкан (Leubner-Metzger and Meins, 1999), в то время как Gh28 и Gh29 представляют пять из 17 семейств белков PR растений (Minic, 2008).Члены семейства Gh2 вовлечены в активацию защитных соединений посредством удаления β-глюкозида (Poulton, 1990; Duroux et al., 1998). Метилэстеразы пектина модифицируют статус этерификации пектина в стенке, влияя на восприимчивость барьера клеточной стенки к грибковым и бактериальным CWDE (Collmer and Keen, 1986). Деэстерификация пектина также влияет на пористость плазмодесм, что может изменить распространение сигнальных молекул во время защитной реакции (Chen et al., 2000).
Роль семейства гликозилтрансфераз CAZy во время защитной реакции менее изучена, чем роль гидролитических ферментов. Примечательно, что существует пять семейств CAZy GT, которые в среднем активируются при абиотических и биотических стрессах у Arabidopsis и ячменя, включая GT1, GT8, GT61, GT75 и GT92. Семейство GT1 включает большое количество генов с широким спектром предполагаемых функций, включая активность UDP-глюкуронозилтрансферазы. Перенося сахара в широкий спектр вторичных метаболитов, UGT повышают стабильность и растворимость агликонов и, следовательно, изменяют их биологическую активность и эффективность в качестве регуляторов защитного ответа (Lim and Bowles, 2004; Langlois-Meurinne et al., 2005). Семейство GT8 катализирует перенос различных сахаров (Glc, Gal, GlcNAC, GalA) на акцепторы липоолигосахаридов, белков, инозитола, олигосахаридов или полисахаридов с использованием нуклеотидных сахарных субстратов (Yin et al., 2011). Члены семейства участвуют в нескольких различных функциях, включая синтез пектинов и ксилана, а также семейство олигосахаридов рафинозы, которые играют роль в реакции на стресс (Kim et al., 2008). На сегодняшний день не сообщалось о том, что семейства GT61, GT75 и GT92 участвуют в защитной реакции растений.Члены семейства GT61 охарактеризовали функции переноса замен арабинозы и ксилозы на 1,4-β-ксилановый бэкон (Anders et al., 2012). Члены GT75 аннотированы как мутазы UDP-Ara (UAM), участвующие в превращении UDP-арабинопиранозы в UDP-арабинофуранозу, что необходимо для образования субстрата UDP-Ara f для арабиноксилана, белка арабиногалактана и пектинового полисахарида.
Перенося сахара в широкий спектр вторичных метаболитов, UGT повышают стабильность и растворимость агликонов и, следовательно, изменяют их биологическую активность и эффективность в качестве регуляторов защитного ответа (Lim and Bowles, 2004; Langlois-Meurinne et al., 2005). Семейство GT8 катализирует перенос различных сахаров (Glc, Gal, GlcNAC, GalA) на акцепторы липоолигосахаридов, белков, инозитола, олигосахаридов или полисахаридов с использованием нуклеотидных сахарных субстратов (Yin et al., 2011). Члены семейства участвуют в нескольких различных функциях, включая синтез пектинов и ксилана, а также семейство олигосахаридов рафинозы, которые играют роль в реакции на стресс (Kim et al., 2008). На сегодняшний день не сообщалось о том, что семейства GT61, GT75 и GT92 участвуют в защитной реакции растений.Члены семейства GT61 охарактеризовали функции переноса замен арабинозы и ксилозы на 1,4-β-ксилановый бэкон (Anders et al., 2012). Члены GT75 аннотированы как мутазы UDP-Ara (UAM), участвующие в превращении UDP-арабинопиранозы в UDP-арабинофуранозу, что необходимо для образования субстрата UDP-Ara f для арабиноксилана, белка арабиногалактана и пектинового полисахарида. биосинтез (Hsieh et al., 2015). С недавним обнаружением арабиноксилана в сосочках ячменя в ответ на попытку проникновения Blumeria graminis f.сп. hordei (Chowdhury et al., 2014), заманчиво предположить, что члены семейства GT61 и GT75 широко вовлечены в защитные реакции. Члены семейства GT92 играют роль в синтезе 1,4-β-галактана (Liwanag et al., 2012), которого относительно много в растянутой древесине, образующейся в ответ на механическое напряжение (Andersson-Gunnerås et al., 2006). . Поэтому, хотя этот общий метаанализ семейств CAZy во время абиотического и биотического стресса не принимает во внимание различия в активности отдельных членов семейства, количестве копий генов или тканеспецифических паттернах экспрессии, он идентифицирует набор семейств CAZy, которые хорошо охарактеризованы с точки зрения реакции на стресс (т.g., Gh27, Gh28), а также менее охарактеризованные (GT61, GT75).
биосинтез (Hsieh et al., 2015). С недавним обнаружением арабиноксилана в сосочках ячменя в ответ на попытку проникновения Blumeria graminis f.сп. hordei (Chowdhury et al., 2014), заманчиво предположить, что члены семейства GT61 и GT75 широко вовлечены в защитные реакции. Члены семейства GT92 играют роль в синтезе 1,4-β-галактана (Liwanag et al., 2012), которого относительно много в растянутой древесине, образующейся в ответ на механическое напряжение (Andersson-Gunnerås et al., 2006). . Поэтому, хотя этот общий метаанализ семейств CAZy во время абиотического и биотического стресса не принимает во внимание различия в активности отдельных членов семейства, количестве копий генов или тканеспецифических паттернах экспрессии, он идентифицирует набор семейств CAZy, которые хорошо охарактеризованы с точки зрения реакции на стресс (т.g., Gh27, Gh28), а также менее охарактеризованные (GT61, GT75).
Вопрос о том, оказывают ли члены этих семейств CAZy специфические или сходные эффекты на мишени клеточной стенки, можно определить, охарактеризовав функцию лежащих в их основе генов. Например, повышенная экспрессия семейств GT8 и GT61 подчеркивает потенциальную роль синтеза пектина и ксилана в реакции растений на стресс у обоих видов. Однако эти семейства содержат членов, которые участвуют во многих различных процессах, и важно более подробно оценить экспрессию и функцию каждого гена.На рис. 3 показаны уровни экспрессии каждого гена из семейства генов GT8 арабидопсиса (рис. 3А) и ячменя (рис. 3В). Большинство генов арабидопсиса GT8 активируются в ответ по крайней мере на один стресс, но, по-видимому, существуют подгруппы, которые реагируют на специфические стрессы. И наоборот, семейство GT8 ячменя разделено на две группы: одна содержит гены, которые не меняются или подавляются в ответ на стресс, а другая содержит гены, которые активируются большинством стрессов.Сравнение генов GT8 чувствительного к стрессу ячменя с охарактеризованными членами семейства GT8 от Arabidopsis (рис. 3C) позволяет предположить, что общие гены чувствительного к стрессу ячменя не ограничиваются кладами с единственной предполагаемой функцией, а распределены между галактинолсинтазой (GolS), ксилановой активности глюкуронозилтрансферазы (GUX) и галактуронозилтрансферазы (GAUT и GATL).
Например, повышенная экспрессия семейств GT8 и GT61 подчеркивает потенциальную роль синтеза пектина и ксилана в реакции растений на стресс у обоих видов. Однако эти семейства содержат членов, которые участвуют во многих различных процессах, и важно более подробно оценить экспрессию и функцию каждого гена.На рис. 3 показаны уровни экспрессии каждого гена из семейства генов GT8 арабидопсиса (рис. 3А) и ячменя (рис. 3В). Большинство генов арабидопсиса GT8 активируются в ответ по крайней мере на один стресс, но, по-видимому, существуют подгруппы, которые реагируют на специфические стрессы. И наоборот, семейство GT8 ячменя разделено на две группы: одна содержит гены, которые не меняются или подавляются в ответ на стресс, а другая содержит гены, которые активируются большинством стрессов.Сравнение генов GT8 чувствительного к стрессу ячменя с охарактеризованными членами семейства GT8 от Arabidopsis (рис. 3C) позволяет предположить, что общие гены чувствительного к стрессу ячменя не ограничиваются кладами с единственной предполагаемой функцией, а распределены между галактинолсинтазой (GolS), ксилановой активности глюкуронозилтрансферазы (GUX) и галактуронозилтрансферазы (GAUT и GATL). На рисунке 4 показаны уровни экспрессии каждого гена из семейства генов GT61 арабидопсиса (рис. 4A) и семейства генов GT61 ячменя (рис. 4B).Нет четкой кластеризации генов, чувствительных и не реагирующих на стресс, как это наблюдается для семейства GT8, при этом гены GT61 активируются при ряде различных стрессов как у арабидопсиса, так и у ячменя. Сравнение чувствительных к стрессу генов ячменя и арабидопсиса GT61 с другими членами семейства GT61, которые были функционально охарактеризованы (рис. 4C), показывает, что чувствительные к стрессу гены не ограничиваются кладами с единственной предполагаемой функцией, т. е. β-(1,2)- активности ксилозилтрансферазы (XylT), ксиланксилозилтрансферазы (XXT) и ксиланарабинофуранозилтрансферазы (XAT).Это еще раз подчеркивает необходимость дальнейшей характеристики генов, связанных с клеточной стенкой, в реакциях на стресс. Сохраняющиеся изменения в экспрессии гена CAZy у разных видов могут указывать на то, что родственные гены рекрутируются для действия на сходные субстраты во время стрессовых реакций.
На рисунке 4 показаны уровни экспрессии каждого гена из семейства генов GT61 арабидопсиса (рис. 4A) и семейства генов GT61 ячменя (рис. 4B).Нет четкой кластеризации генов, чувствительных и не реагирующих на стресс, как это наблюдается для семейства GT8, при этом гены GT61 активируются при ряде различных стрессов как у арабидопсиса, так и у ячменя. Сравнение чувствительных к стрессу генов ячменя и арабидопсиса GT61 с другими членами семейства GT61, которые были функционально охарактеризованы (рис. 4C), показывает, что чувствительные к стрессу гены не ограничиваются кладами с единственной предполагаемой функцией, т. е. β-(1,2)- активности ксилозилтрансферазы (XylT), ксиланксилозилтрансферазы (XXT) и ксиланарабинофуранозилтрансферазы (XAT).Это еще раз подчеркивает необходимость дальнейшей характеристики генов, связанных с клеточной стенкой, в реакциях на стресс. Сохраняющиеся изменения в экспрессии гена CAZy у разных видов могут указывать на то, что родственные гены рекрутируются для действия на сходные субстраты во время стрессовых реакций. Альтернативно, гены из одного и того же семейства могут быть рекрутированы для модификации разных субстратов, но сходным образом.
Альтернативно, гены из одного и того же семейства могут быть рекрутированы для модификации разных субстратов, но сходным образом.
Рисунок 3. Анализ представителей семейства GT8 после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) .Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1. Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. (C) Филогенетическое древо членов семейства GT8 из арабидопсиса и ячменя с предполагаемыми функциями, присвоенными каждой кладе. Красными точками отмечены гены ячменя, которые активируются в ответ на стресс (B) .
Красными точками отмечены гены ячменя, которые активируются в ответ на стресс (B) .
Рисунок 4. Анализ членов семейства GT61 после абиотических и биотических стрессов у арабидопсиса (A) и ячменя (B) . Обилие транскриптов определяли путем метаанализа наборов данных микрочипов, собранных из базы данных экспрессии растений (PLEXdb; Dash et al., 2012), с использованием экспериментов, перечисленных в таблице 2. Значения показывают среднее логарифмическое (2)-кратное увеличение индукции для представителей каждого Семейство генов CAZy присутствует на массиве генома ATh2 Arabidopsis Affymetrix 22K ATh2 и на генетическом чипе 22K Barley1.Иерархическая кластеризация была выполнена на основе коэффициентов корреляции Пирсона для каждого набора данных и семейства CAZy. (C) Филогенетическое древо членов семейства GT61 из арабидопсиса, ячменя и риса с предполагаемыми функциями, назначенными для каждой клады.
Перспективы и резюме
Основой для этого обзора было рассмотрение активности, связанной с клеточной стенкой и полисахаридами, которая влияет на реакцию на биотический и абиотический стресс, и выделение тех, которые могут выполнять общую функцию, стимулируя ремоделирование клеточной стенки в качестве прямого ответа на абиотический стресс или атаку патогенов. .Генетические и трансгенные данные свидетельствуют о том, что модификация активности специфических клеточных стенок оказывает выраженное влияние на толерантность к стрессу. В некоторых случаях сходные семейства генов, по-видимому, модулируют эффект различных биотических и абиотических стрессов внутри и между разными видами, что подразумевает, что общие механизмы могли быть задействованы для воздействия на, казалось бы, несопоставимые типы стресса. Это подтверждается более широким анализом всего транскриптома, который указывает на сходные ответы отдельных генов, связанных с клеточной стенкой, и даже семейств CAZy на различные абиотические и биотические стрессы.Приводят ли эти перекрытия в экспрессии генов к аналогичным изменениям в структуре клеточных стенок, в большинстве случаев еще предстоит подтвердить, особенно в случае пектинов и ксиланов, которые демонстрируют явные различия в количестве между моделями однодольных и двудольных растений. В самом деле, функции многих генов, связанных с клеточной стенкой, еще не описаны во время нормального роста и развития, не говоря уже о реакциях на стресс.
.Генетические и трансгенные данные свидетельствуют о том, что модификация активности специфических клеточных стенок оказывает выраженное влияние на толерантность к стрессу. В некоторых случаях сходные семейства генов, по-видимому, модулируют эффект различных биотических и абиотических стрессов внутри и между разными видами, что подразумевает, что общие механизмы могли быть задействованы для воздействия на, казалось бы, несопоставимые типы стресса. Это подтверждается более широким анализом всего транскриптома, который указывает на сходные ответы отдельных генов, связанных с клеточной стенкой, и даже семейств CAZy на различные абиотические и биотические стрессы.Приводят ли эти перекрытия в экспрессии генов к аналогичным изменениям в структуре клеточных стенок, в большинстве случаев еще предстоит подтвердить, особенно в случае пектинов и ксиланов, которые демонстрируют явные различия в количестве между моделями однодольных и двудольных растений. В самом деле, функции многих генов, связанных с клеточной стенкой, еще не описаны во время нормального роста и развития, не говоря уже о реакциях на стресс. Это подчеркивает необходимость дальнейшего распространения технологий редактирования генома на целые семейства CAZy и разработки высокопроизводительных методологий химического анализа клеточных стенок, способных быть нацеленными на отдельные типы клеток.
Это подчеркивает необходимость дальнейшего распространения технологий редактирования генома на целые семейства CAZy и разработки высокопроизводительных методологий химического анализа клеточных стенок, способных быть нацеленными на отдельные типы клеток.
Вклад авторов
KH, AL, MT, NS и JC задумали этот обзор, составили и отредактировали рукопись, утвердили окончательную версию перед публикацией и согласились нести ответственность за все аспекты работы, гарантируя, что вопросы, связанные с точностью или целостностью любая часть работы должным образом исследована и решена.
Финансирование
KH выражает благодарность исследовательской программе шотландского правительства. AL, NS и JC были поддержаны грантами Австралийского исследовательского совета.MT был поддержан ARC Future Fellowship.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Ссылки
Абебе Т., Мелмаи К., Берг В. и Уайз Р. П. (2010). Реакция колоса ячменя на засуху: экспрессия генов в чешуе, цветковом чешуе, ости и семени. Функц. интегр. Геномика 10, 191–205.doi: 10.1007/s10142-009-0149-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Aditya, J., Lewis, J., Shirley, N.J., Tan, H.T., Henderson, M., Fincher, G.B., et al. (2015). Динамика заражения злаковыми цистообразными нематодами различается между чувствительными и устойчивыми сортами ячменя и приводит к изменениям уровня (1, 3; 1, 4)-β-глюкана и количества транскриптов гена HvCslF. Новый фитол . 207, 135–147. doi: 10.1111/nph.13349
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аист, Дж.Р. и Исраэль, Х.В. (1977). Формирование сосочков: сроки и значение при проникновении Erysiphe graminis hordei в колеоптилы ячменя. Фитопатология 67, 455–461. doi: 10.1094/Фито-67-455
Полнотекстовая перекрестная ссылка | Академия Google
Альбершейм, П. , Дарвилл, А., Робертс, К., Седерофф, Р., и Штехелин, А. (2011). Клеточные стенки и взаимодействие растений и микробов: Garland Science . Нью-Йорк, штат Нью-Йорк: Тейлор и Фрэнсис.
, Дарвилл, А., Робертс, К., Седерофф, Р., и Штехелин, А. (2011). Клеточные стенки и взаимодействие растений и микробов: Garland Science . Нью-Йорк, штат Нью-Йорк: Тейлор и Фрэнсис.
Ан, С. Х., Зон, К.Х., Чой, Х.В., Хван, И.С., Ли, С.К., и Хван, Б.К. (2008). Ингибитор метилэстеразы пектина перца белок CaPMEI1 необходим для противогрибковой активности, устойчивости к базальным заболеваниям и толерантности к абиотическому стрессу. Планта 228, 61–78. doi: 10.1007/s00425-008-0719-z
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Андерс, Н., Уилкинсон, М.Д., Лавгроув, А., Фриман, Дж., Трифона, Т., Пеллни, Т.К., и соавт. (2012). Гликозилтрансферазы семейства 61 опосредуют перенос арабинофуранозила на ксилан в травах. Проц. Натл. акад. науч. США 109, 989–993. doi: 10.1073/pnas.1115858109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Andersson-Gunneras, S., Mellerowicz, E. J., Love, J., Segerman, B., Ohmiya, Y., Coutinho, P.M., et al. (2006). Биосинтез обогащенной целлюлозой натянутой древесины у Populus: глобальный анализ транскриптов и метаболитов идентифицирует биохимические регуляторы и регуляторы развития в биосинтезе вторичной стенки. Завод Ж. 45, 144–165.doi: 10.1111/j.1365-313X.2005.02584.x
J., Love, J., Segerman, B., Ohmiya, Y., Coutinho, P.M., et al. (2006). Биосинтез обогащенной целлюлозой натянутой древесины у Populus: глобальный анализ транскриптов и метаболитов идентифицирует биохимические регуляторы и регуляторы развития в биосинтезе вторичной стенки. Завод Ж. 45, 144–165.doi: 10.1111/j.1365-313X.2005.02584.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Аткинсон, Нью-Джерси, Лилли, С.Дж., и Урвин, П.Е. (2013). Выявление генов, участвующих в реакции арабидопсиса на одновременные биотические и абиотические стрессы. Завод физиол. 162, 2028–2041 гг. doi: 10.1104/стр.113.222372
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Беллинкампи Д., Червоне Ф. и Лионетти В. (2014). Динамика клеточных стенок растений и связанная со стенками восприимчивость к взаимодействиям растений и патогенов. Перед. Растениевод. 5:228. doi: 10.3389/fpls.2014.00228
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бхаттараи К. К., Атамиан Х.С., Калошян И. и Юлгем Т. (2010). Факторы транскрипции типа WRKY72 вносят вклад в базальный иммунитет у томата и арабидопсиса, а также в резистентность между генами, опосредованную геном R томата Mi-1. Завод J. 63, 229–240. doi: 10.1111/j.1365-313X.2010.04232.x
К., Атамиан Х.С., Калошян И. и Юлгем Т. (2010). Факторы транскрипции типа WRKY72 вносят вклад в базальный иммунитет у томата и арабидопсиса, а также в резистентность между генами, опосредованную геном R томата Mi-1. Завод J. 63, 229–240. doi: 10.1111/j.1365-313X.2010.04232.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бхуян, Н.Х., Селварадж Г., Вей Ю. и Кинг Дж. (2009). Профилирование экспрессии генов и замалчивание показывают, что биосинтез монолигнола играет критическую роль в защите пшеницы от проникновения мучнистой росы. Дж. Экспл. Бот. 60, 509–521. doi: 10.1093/jxb/ern290
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бодд, Дж., Чо, С., Крюгер, В. М., и Мюльбауэр, Г. Дж. (2006). Транскриптомный анализ взаимодействия ячменя и Fusarium graminearum. Мол.Взаимодействие растительных микробов. 19, 407–417. doi: 10.1094/MPMI-19-0407
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Боллер, Т. , и Феликс, Г. (2009). Возрождение элиситоров: восприятие связанных с микробами молекулярных паттернов и сигналов опасности рецепторами распознавания паттернов. год. Преподобный завод биол. 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
, и Феликс, Г. (2009). Возрождение элиситоров: восприятие связанных с микробами молекулярных паттернов и сигналов опасности рецепторами распознавания паттернов. год. Преподобный завод биол. 379–406. doi: 10.1146/annurev.arplant.57.032905.105346
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бертон, Р.А., Гидли, М.Дж., и Финчер, Г.Б. (2010). Неоднородность химического состава, строения и функции клеточных стенок растений. Нац. хим. биол. 6, 724–732. doi: 10.1038/nchembio.439
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бертон, Р. А., Уилсон, С. М., Хрмова, М., Харви, А. Дж., Ширли, Н. Дж., Медхерст, А., и др. (2006). Целлюлозосинтазоподобные гены CslF опосредуют синтез (1, 3; 1, 4)-β-D-глюканов клеточной стенки. Наука 311, 1940–1942 гг.doi: 10.1126/наука.1122975
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кэл, А. Дж., Лю, Д., Молеон, Р. , Син, Ю. И. К., и Серрадж, Р. (2013). Транскриптомное профилирование зоны удлинения листьев в условиях засухи у контрастных сортов риса. PLoS ONE 8:e54537. doi: 10.1371/journal.pone.0054537
, Син, Ю. И. К., и Серрадж, Р. (2013). Транскриптомное профилирование зоны удлинения листьев в условиях засухи у контрастных сортов риса. PLoS ONE 8:e54537. doi: 10.1371/journal.pone.0054537
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Каньо-Дельгадо, А., Пенфилд, С., Смит, К., Кэтли, М., и Беван, М. (2003).Снижение синтеза целлюлозы вызывает лигнификацию и защитные реакции у Arabidopsis thaliana . Завод J. 34, 351–362. doi: 10.1046/j.1365-313X.2003.01729.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чен, М. Х., Шэн, Дж., Хинд, Г., Ханда, А. К., и Цитовски, В. (2000). Взаимодействие между белком перемещения вируса табачной мозаики и пектинметилэстеразами клетки-хозяина необходимо для перемещения вируса от клетки к клетке. ЭМБО Дж. 19, 913–920. doi: 10.1093/emboj/19.5.913
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Chen, Z. , Hong, X., Zhang, H., Wang, Y., Li, X., Zhu, J.K., et al. (2005). Нарушение гена синтазы целлюлозы, AtCesA8/IRX1, повышает устойчивость арабидопсиса к засухе и осмотическому стрессу. Завод J. 43, 273–283. doi: 10.1111/j.1365-313X.2005.02452.x
, Hong, X., Zhang, H., Wang, Y., Li, X., Zhu, J.K., et al. (2005). Нарушение гена синтазы целлюлозы, AtCesA8/IRX1, повышает устойчивость арабидопсиса к засухе и осмотическому стрессу. Завод J. 43, 273–283. doi: 10.1111/j.1365-313X.2005.02452.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чо, С.К., Ким, Дж.Э., Парк, Дж. А., Эом, Т. Дж., и Ким, В. Т. (2006). Конститутивная экспрессия индуцируемого абиотическим стрессом острого перца CaXTh4 , который кодирует гомолог ксилоглюкан-эндотрансглюкозилазы/гидролазы, улучшает засухоустойчивость и солеустойчивость у трансгенных растений арабидопсиса. Письмо ФЭБС. 580, 3136–3144. doi: 10.1016/j.febslet.2006.04.062
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чой, Дж. Ю., Сео, Ю. С., Ким, С. Дж., Ким, В. Т., и Шин, Дж. С. (2011).Конститутивная экспрессия CaXTh4, ксилоглюкановой эндотрансглюкозилазы/гидролазы острого перца, повышала устойчивость растений томата к солевым и засушливым стрессам без фенотипических дефектов ( Solanum lycopersicum сорта Dotaerang). Представитель растительных клеток 30, 867–877. doi: 10.1007/s00299-010-0989-3
Представитель растительных клеток 30, 867–877. doi: 10.1007/s00299-010-0989-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чоудхури Дж., Хендерсон М., Швейцер П., Бертон Р. А., Финчер Г. Б. и Литтл А. (2014). Дифференциальное накопление каллозы, арабиноксилана и целлюлозы в непроникающих и проникающих сосочках на листьях ячменя, инфицированных Blumeria graminis f.сп. хорд. Новый Фитол. 204, 650–660. doi: 10.1111/nph.12974
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Колбрук, Э. Х., Крейссен, Г., МакГранн, Г. Р. Д., Дреос, Р., Лэмб, К., и Бойд, Л. А. (2012). Приобретенная устойчивость широкого спектра действия у ячменя, индуцированная патосистемой Pseudomonas, имеет общие компоненты транскрипции с системной приобретенной устойчивостью Arabidopsis. Мол. Взаимодействие растительных микробов. 25, 658–667. doi: 10.1094/MPMI-09-11-0246
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коллмер, Н. и Кин Т. (1986). Роль пектиновых ферментов в патогенезе растений. год. Преподобный Фитопат. 24, 383–409. doi: 10.1146/annurev.py.24.0
и Кин Т. (1986). Роль пектиновых ферментов в патогенезе растений. год. Преподобный Фитопат. 24, 383–409. doi: 10.1146/annurev.py.24.0
.002123
Полнотекстовая перекрестная ссылка | Академия Google
Коминелли Э., Сала Т., Кальви Д., Гусмароли Г. и Тонелли К. (2008). Сверхэкспрессия гена AtMYB41 Arabidopsis изменяет рост клеток и проницаемость поверхности листа. Завод Ж. 53, 53–64. doi: 10.1111/j.1365-313X.2007.03310.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кулен, С., Proietti, S., Hickman, R., Davila Olivas, N.H., Huang, P.-P., Van Verk, M.C., et al. (2016). Динамика транскриптома арабидопсиса при последовательных биотических и абиотических стрессах. Завод J. 86, 249–267. doi: 10.1111/tpj.13167
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Craigon, D.J., James, N., Okyere, J., Higgins, J., Jotham, J., and May, S. (2004). NASCArrays: репозиторий данных микрочипов, созданных службой транскриптомики NASC. Рез. нуклеиновых кислот. 32, Д575–Д577. doi: 10.1093/nar/gkh233
Рез. нуклеиновых кислот. 32, Д575–Д577. doi: 10.1093/nar/gkh233
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Cu, S., Collins, H.M., Betts, N.S., March, T.J., Janusz, A., Stewart, D.C., et al. (2016). Поглощение воды зерном ячменя: физиология; генетика и промышленное применение. Растениеводство . 242, 260–269. doi: 10.1016/j.plantsci.2015.08.009
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Катлер, Дж. М., Шахан, К. В., и Степонкус, П.Л. (1980). Влияние водного дефицита и осмотической адаптации на удлинение листьев риса. Растениеводство. 20, 314–318. doi: 10.2135/cropsci1980.0011183X002000030006x
Полнотекстовая перекрестная ссылка | Академия Google
Дэш, С., Ван Хемерт, Дж., Хонг, Л., Уайз, Р.П., и Дикерсон, Дж.А. (2012). PLEXdb: ресурсы экспрессии генов для растений и фитопатогенов. Рез. нуклеиновых кислот. 40, Д1194–Д1201. doi: 10.1093/nar/gkr938
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дэвис, Л. J., и Urwin, PE (2012). Клеточные стенки синцитиев, образованные Heterodera schachtii в Arabidopsis thaliana , богаты метилэтерифицированным пектином. Сигнал завода. Поведение 7, 1404–1406. doi: 10.4161/psb.21925
J., и Urwin, PE (2012). Клеточные стенки синцитиев, образованные Heterodera schachtii в Arabidopsis thaliana , богаты метилэтерифицированным пектином. Сигнал завода. Поведение 7, 1404–1406. doi: 10.4161/psb.21925
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Де Вос, М., Ван Остен, В.Р., Ван Поке, Р.М., Ван Пелт, Дж.А., Позо, М.Дж., Мюллер, М.Дж., и соавт. (2005). Изменения сигнальной сигнатуры и транскриптома арабидопсиса во время атаки патогенов и насекомых. Мол. Взаимодействие растительных микробов. 18, 923–937. doi: 10.1094/MPMI-18-0923
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дельгадо-Сересо, М., Санчес-Родригес, К., Эскудеро, В., Миедес, Э., Фернандес, П.В., Хорда, Л., и др. (2012). Гетеротримерный G-белок арабидопсиса регулирует защиту клеточной стенки и устойчивость к некротрофным грибам. Мол. Растение. 5, 98–114. дои: 10.1093/mp/ssr082
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дельп, Г. , Градин, Т., Ахман, И., и Йонссон, Л.М. (2009). Микроматричный анализ взаимодействия между тлей Rhopalosiphum padi и растениями-хозяевами выявляет как различия, так и сходства между восприимчивыми и частично устойчивыми линиями ячменя. Мол. Жене. Геномика 281, 233–248. doi: 10.1007/s00438-008-0409-3
, Градин, Т., Ахман, И., и Йонссон, Л.М. (2009). Микроматричный анализ взаимодействия между тлей Rhopalosiphum padi и растениями-хозяевами выявляет как различия, так и сходства между восприимчивыми и частично устойчивыми линиями ячменя. Мол. Жене. Геномика 281, 233–248. doi: 10.1007/s00438-008-0409-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Денансе, Н., Раноча, П., Ориа, Н., Барле, X., Ривьер, М.П., Ядета, К.А., и соавт.(2013). Опосредованная Arabidopsis wat1 ( стенки тонкие1 ) устойчивость к бактериальному сосудистому возбудителю Ralstonia solanacearum сопровождается перекрестной регуляцией метаболизма салициловой кислоты и триптофана. Завод J. 73, 225–239. doi: 10.1111/tpj.12027
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Deubert, K.H., and Rohde, R.A. (1971). «Ферменты нематод», в Паразитические нематоды растений: цитогенетика, взаимодействия хозяин-паразит и физиология , Vol. 2, ред. Б. М. Цукерман, В. Ф. Май и Р. А. Роде (Нью-Йорк, штат Нью-Йорк: Academic Press), 73–90.
2, ред. Б. М. Цукерман, В. Ф. Май и Р. А. Роде (Нью-Йорк, штат Нью-Йорк: Academic Press), 73–90.
Академия Google
Дюру, Л., Дельмотт, Ф.М., Ланселин, Ж.-М., Керавис, Г., и Джей-Аллеанд, К. (1998). Взгляд на метаболизм нафтохинона: катализируемый β-глюкозидазой гидролиз гидроюглона β-D-глюкопиранозида. Биохим. J. 333, 275–283. дои: 10.1042/bj3330275
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эйзен, М. Б., Спеллман, П.Т., Браун П.О. и Ботштейн Д. (1998). Кластерный анализ и отображение полногеномных паттернов экспрессии. Проц. Натл. акад. науч. США 95, 14863–14868. doi: 10.1073/pnas.95.25.14863
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ellinger, D., Naumann, M., Falter, C., Zwikowics, C., Jamrow, T., Manisseri, C., et al. (2013). Повышенное раннее отложение каллозы приводит к полной устойчивости арабидопсиса к проникновению мучнистой росы. Завод физиол. 161, 1433–1444. doi: 10.1104/стр.112.211011
doi: 10.1104/стр.112.211011
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эллис, К., и Тернер, Дж. Г. (2001). Мутант арабидопсиса cev1 имеет конститутивно активные жасмонатные и этиленовые сигнальные пути и повышенную устойчивость к патогенам. Растительная клетка 13, 1025–1033. doi: 10.1105/tpc.13.5.1025
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фэн Л., Линкер Р., Гепштейн С., Танимото, Э., Ямамото, Р., и Нойманн, П.М. (2006). Прогрессирующее ингибирование водным дефицитом растяжимости и роста клеточных стенок вдоль зоны растяжения корней кукурузы связано с повышенным метаболизмом лигнина и прогрессирующим накоплением фенольных смол в стенках. Завод физиол. 140, 603–612. doi: 10.1104/стр.105.073130
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Феррари С., Саватин Д. В., Сицилия Ф., Граменья Г., Червоне Ф. и Де Лоренцо Г.(2013). Олигогалактурониды: молекулярные паттерны, связанные с повреждением растений, и регуляторы роста и развития. Перед. Растениевод. 4:49. doi: 10.3389/fpls.2013.00049
Перед. Растениевод. 4:49. doi: 10.3389/fpls.2013.00049
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрай, С.К., Смит, Р.К., Ренвик, К.Ф., Мартин, Д.Дж., Ходж, С.К., и Мэтьюз, К.Дж. (1992). Ксилоглюкан эндотрансгликозилаза, новый фермент, разрыхляющий стенки растений. Биохим. Дж. 282, 821–828. дои: 10.1042/bj2820821
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гехан, М.А., Гринхэм, К., Моклер, Т.С., и МакКланг, Ч.Р. (2015). Транскрипционные сети — урожай, часы и абиотический стресс. Курс. мнение Растение Биол . 24, 39–46. doi: 10.1016/j.pbi.2015.01.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гонсалес-Перес, С., Гутьеррес, Дж., Гарсия-Гарсия, Ф., Осуна, Д., Допазо, Дж., Лоренцо, О., и др. (2011). Ранние защитные реакции транскрипции в культуре суспензии клеток арабидопсиса в условиях яркого света. Завод физиол. 156, 1439–1456. doi: 10.1104/стр.111.177766
doi: 10.1104/стр.111.177766
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Харб, А., Кришнан, А., Амбаварам, М.М., и Перейра, А. (2010). Молекулярно-физиологический анализ стресса от засухи у арабидопсиса выявляет ранние реакции, ведущие к акклиматизации роста растений. Завод физиол. 154, 1254–1271. doi: 10.1104/стр.110.161752
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эрнандес-Бланко, К., Feng, D.X., Hu, J., Sánchez-Vallet, A., Deslandes, L., Llorente, F., et al. (2007). Нарушение синтазы целлюлозы, необходимой для образования вторичной клеточной стенки арабидопсиса, повышает устойчивость к болезням. Растительная клетка 19, 890–903. doi: 10.1105/tpc.106.048058
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хофманн, Дж., Юссеф-Банора, М., де Алмейда-Энглер, Дж., и Грундлер, Ф.М. (2010). Роль отложения каллозы вдоль плазмодесм в местах питания нематод. Мол. Взаимодействие растительных микробов. 23, 549–557. doi: 10.1094/MPMI-23-5-0549
Мол. Взаимодействие растительных микробов. 23, 549–557. doi: 10.1094/MPMI-23-5-0549
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хок С., Аттард А. и Келлер Х. (2010). Получение максимума от хозяина: как патогены заставляют растения сотрудничать при болезни. Мол. Взаимодействие растительных микробов. 23, 1253–1259. doi: 10.1094/MPMI-04-10-0103
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Се, Ю. С., Чжан, К., Яп, К., Ширли, Н.Дж., Ланштайн Дж., Нельсон С.Дж. и соавт. (2015). Генетика, профили транскрипции и каталитические свойства семейства udp-арабинозомутазы из ячменя. Биохимия 55, 322–334. doi: 10.1021/acs.biochem.5b01055
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Иноуэ, С., Аист, Дж. Р., и Мако, В. (1994). Более раннее образование сосочков и устойчивость к мучнистой росе ячменя, индуцированные экстрактом, регулирующим сосочков. Физиол. Мол. Завод Патол. 44, 433–440. doi: 10.1016/S0885-5765(05)80099-2
Физиол. Мол. Завод Патол. 44, 433–440. doi: 10.1016/S0885-5765(05)80099-2
Полнотекстовая перекрестная ссылка | Академия Google
Джейкобс А.К., Липка В., Бертон Р.А., Панструга Р., Стрижов Н., Шульце-Леферт П. и соавт. (2003). Синтаза каллозы Arabidopsis, GSL5 , необходима для образования мозолей в ранах и сосочках. Растительная клетка. 15, 2503–2513. doi: 10.1105/tpc.016097
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дженсен, М. К., Хагедорн, П.H., Torres-Zabala, D., Grant, M.R., Rung, J.H., Collinge, D.B., et al. (2008). Регуляция транскрипции фактором транскрипции NAC (NAM-ATAF1, 2-CUC2) ослабляет передачу сигналов ABA для эффективной базальной защиты от Blumeria graminis f.sp. hordei у арабидопсиса. Завод J. 56, 867–880. doi: 10.1111/j.1365-313X.2008.03646.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кемпема, Л. А., Куи, X., Хольцер, Ф. М., и Уоллинг, Л. Л.(2007). Транскриптом арабидопсиса изменяется в ответ на питание нимф белокрылки серебристолистной флоэмой. сходства и различия в реакциях на тлю. Завод физиол. 143, 849–865. doi: 10.1104/стр.106.0
М., и Уоллинг, Л. Л.(2007). Транскриптом арабидопсиса изменяется в ответ на питание нимф белокрылки серебристолистной флоэмой. сходства и различия в реакциях на тлю. Завод физиол. 143, 849–865. doi: 10.1104/стр.106.0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ким, М.С., Чо, С.М., Канг, Э.Ю., Им, Ю.Дж., Хванбо, Х., Ким, Ю.К., и соавт. (2008). Галактинол является сигнальным компонентом индуцированной системной резистентности, вызванной колонизацией корней Pseudomonas chlororaph is O6. Мол. Взаимодействие растительных микробов. 21, 1643–1653. doi: 10.1094/MPMI-21-12-1643
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кимпара Т., Аохара Т., Сога К., Вакабаяси К., Хосон Т., Цумурая Ю. и др. (2008). Активность β-1,3:1,4-глюкансинтазы в проростках риса под водой. Энн. Бот. 102, 221–226. doi: 10.1093/aob/mcn077
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Квон Ю. , Ким С. Х., Jung, M.S., Kim, M.S., Oh, J.E., Ju, H.W., et al. (2007). Arabidopsis hot2 кодирует эндохитиназоподобный белок, необходимый для устойчивости к жаре, соли и засухе. Завод Ж. 49, 184–193. doi: 10.1111/j.1365-313X.2006.02950.x
, Ким С. Х., Jung, M.S., Kim, M.S., Oh, J.E., Ju, H.W., et al. (2007). Arabidopsis hot2 кодирует эндохитиназоподобный белок, необходимый для устойчивости к жаре, соли и засухе. Завод Ж. 49, 184–193. doi: 10.1111/j.1365-313X.2006.02950.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Langlois-Meurinne, M., Gachon, C.M.M., and Saindrean, P. (2005). Патоген-чувствительная экспрессия генов гликозилтрансфераз UGT73B3 и UGT73B5 необходима для устойчивости к Pseudomonas syringae pv томата у Arabidopsis. Завод физиол. 139, 1890–1901 гг. doi: 10.1104/стр.105.067223
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ле Галль, Х., Филипп, Ф., Домон, Дж. М., Жилле, Ф., Пеллу, Дж., и Район, К. (2015). Метаболизм клеточной стенки в ответ на абиотический стресс. Растения 4, 112–166. doi: 10.3390/plants4010112
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лейбнер-Мецгер, Г. , и Мейнс, Ф.Дж. (1999). «Функции и регуляция растительных β-1,3-глюканаз (PR-2)», в «Связанные с патогенезом белки в растениях» , под редакцией S.Датта и С. Мутукришнан (Флорида, Флорида: CRC Press LLC Boca Raton), 49–76.
, и Мейнс, Ф.Дж. (1999). «Функции и регуляция растительных β-1,3-глюканаз (PR-2)», в «Связанные с патогенезом белки в растениях» , под редакцией S.Датта и С. Мутукришнан (Флорида, Флорида: CRC Press LLC Boca Raton), 49–76.
Академия Google
Ли Х., Ян С., Чжао Л., Тан Дж., Чжан К., Гао Ф. и др. (2014). Активация генов, связанных с клеточной стенкой, связанная с ацетилированием гистонов, участвует в набухании корней кукурузы, вызванном солевым стрессом. BMC Растение Биол. 14:105. дои: 10.1186/1471-2229-14-105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лионетти, В., Червоне, Ф.и Беллинкампи, Д. (2012). Метиловая этерификация пектина играет роль во взаимодействиях растений и патогенов и влияет на устойчивость растений к болезням. J. Физиол растений. 169, 1623–1630. doi: 10.1016/j.jplph.2012.05.006
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Liu, H., Ma, Y. , Chen, N.A., Guo, S., Liu, H., Guo, X., et al. (2014). Сверхэкспрессия индуцируемого стрессом OsBURP16 , β-субъединицы полигалактуроназы 1, снижает содержание пектина и адгезию клеток и повышает чувствительность к абиотическому стрессу у риса. Окружающая среда растительных клеток. 37, 1144–1158. doi: 10.1111/pce.12223
, Chen, N.A., Guo, S., Liu, H., Guo, X., et al. (2014). Сверхэкспрессия индуцируемого стрессом OsBURP16 , β-субъединицы полигалактуроназы 1, снижает содержание пектина и адгезию клеток и повышает чувствительность к абиотическому стрессу у риса. Окружающая среда растительных клеток. 37, 1144–1158. doi: 10.1111/pce.12223
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Liwanag, A.J.M., Ebert, B., Verhertbruggen, Y., Rennie, E.A., Rautengarten, C., Oikawa, A., et al. (2012). Биосинтез пектина: GALS1 в Arabidopsis thaliana представляет собой β-1,4-галактан β-1,4-галактозилтрансферазу. Растительная клетка 24, 5024–5036. doi: 10.1105/tpc.112.106625
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ломбард, В., Голаконда Рамулу, Х., Друла, Э., Коутиньо, П.М., и Хенриссат, Б. (2014). База данных углеводно-активных ферментов (CAZy) в 2013 г. Nucleic Acids Res. 42, Д490–Д495. doi: 10.1093/nar/gkt1178
doi: 10.1093/nar/gkt1178
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Manabe, Y., Nafisi, M., Verhertbruggen, Y., Orfila, C., Gille, S., Rautengarten, C., et al. (2011). Мутация с потерей функции REDUCED WALL ACETYLATION2 у Arabidopsis приводит к снижению ацетилирования клеточной стенки и повышению устойчивости к Botrytis cinerea . Завод физиол. 155, 1068–1078. doi: 10.1104/стр.110.168989
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мангельсен, Э., Килиан, Дж., Хартер, К., Янссон, К., Ванке, Д., и Сандберг, Э. (2011). Транскриптомный анализ высокотемпературного стресса в развивающихся зерновках ячменя: ранние реакции на стресс и влияние на биосинтез запасных соединений. Мол. Завод 4, 97–115. doi: 10.1093/mp/ssq058
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
МакГранн, Г.Р., Таунсенд, Б.Дж., Антонив, Дж.Ф., Ашер, М.Дж., и Мутаса-Гёттгенс, Э. С. (2009). Ячмень вызывает аналогичную раннюю базальную защитную реакцию во время взаимодействия хозяина и не хозяина с корневыми паразитами Polymyxa. евро. Дж. Плант Патол. 123, 5–15. doi: 10.1007/s10658-008-9332-z
С. (2009). Ячмень вызывает аналогичную раннюю базальную защитную реакцию во время взаимодействия хозяина и не хозяина с корневыми паразитами Polymyxa. евро. Дж. Плант Патол. 123, 5–15. doi: 10.1007/s10658-008-9332-z
Полнотекстовая перекрестная ссылка | Академия Google
Миллетт Б.П., Сюн Ю., Даль С.К., Штеффенсон Б.Дж. и Мюльбауэр Г.Дж. (2009). Дикий ячмень накапливает разные наборы транскриптов в ответ на патогены с разным трофическим образом жизни. Физиол. Мол. Завод Патол . 74, 91–98. doi: 10.1016/j.pmpp.2009.09.006
Полнотекстовая перекрестная ссылка | Академия Google
Мершбахер, Б., и Мендген, К. (2012). «Структурный аспект защиты растений», в Механизмах устойчивости к болезням растений , под ред. А. Дж. Слюсаренко, Р. С. С. Фрейзер и Л. К. ван Лун (Берлин: Springer Science & Business Media), 231–277.
Академия Google
Мур, Дж. П., Викре-Гибуэн, М., Фаррант, Дж. М.и Дриуич, А. (2008). Адаптация клеточных стенок высших растений к потере воды: засуха против высыхания. Физиол. Завод . 134, 237–245. doi: 10.1111/j.1399-3054.2008.01134.x
Физиол. Завод . 134, 237–245. doi: 10.1111/j.1399-3054.2008.01134.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mouille, G., Robin, S., Lecomte, M., Pagant, S., and Höfte, H. (2003). Классификация и идентификация мутантов клеточной стенки арабидопсиса с помощью микроспектроскопии с преобразованием Фурье в инфракрасном диапазоне (FT-IR). Завод J. 35, 393–404.doi: 10.1046/j.1365-313X.2003.01807.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нафиси, М., Фимогнари, Л., и Сакураги, Ю. (2015). Взаимодействия между клеточной стенкой и фитогормонами при взаимодействии растений и некротрофных патогенов. Фитохимия 112, 63–71. doi: 10.1016/j.phytochem.2014.11.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нисимура, М. Т., Штейн, М., Хоу, Б. Х., Фогель, Дж. П., Эдвардс, Х.и Somerville, Южная Каролина (2003). Потеря синтазы каллозы приводит к устойчивости к салициловой кислоте. Наука 301, 969–972. doi: 10.1126/наука.1086716
Наука 301, 969–972. doi: 10.1126/наука.1086716
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Оз, М. Т., Йилмаз, Р., Эйдоган, Ф., Де Грааф, Л., и Юсел, М. Октем, Х. А. (2009). Микроматричный анализ поздней реакции листьев ячменя ( Hordeum vulgare L.) на токсичность бора. тюрк. Дж. Агрик. За. 33, 191–202.doi: 10.3906/tar-0806-22
Полнотекстовая перекрестная ссылка | Академия Google
Поли М., Гилле С., Лю Л., Мансури Н., де Соуза А., Шултинк А. и др. (2013). Биосинтез гемицеллюлозы. Планта 238, 627–642. doi: 10.1007/s00425-013-1921-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пеллу, Дж., Рустеруччи, К., и Меллерович, Э. Дж. (2007). Новое понимание структуры и функции пектинметилэстеразы. Trends Plant Sci. 12, 267–277. doi: 10.1016/j.tplants.2007.04.001
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Пураник, С. , Саху, П. П., Шривастава, П. С., и Прасад, М. (2012). Белки NAC: регуляция и роль в стрессоустойчивости. Trends Plant Sci. 17, 369–381. doi: 10.1016/j.tplants.2012.02.004
, Саху, П. П., Шривастава, П. С., и Прасад, М. (2012). Белки NAC: регуляция и роль в стрессоустойчивости. Trends Plant Sci. 17, 369–381. doi: 10.1016/j.tplants.2012.02.004
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рай К.М., Баласубраманян В.К., Велкер К.М., Панг М., Хии М.М. и Менду В.(2015). Комплексный анализ всего генома и разработка веб-ресурсов о ферментах, разрушающих клеточную стенку фитопаразитических нематод. BMC Растение Биол. 15:187. doi: 10.1186/s12870-015-0576-4
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Раксеги М., Лавгроув А., Балла К., Ланг Л., Бедо З., Вейс О. и др. (2014). Влияние теплового и засушливого стресса на структуру и состав арабиноксилана и β-глюкана в зерне пшеницы. Углеводы Полим. 102, 557–565. doi: 10.1016/j.carbpol.2013.12.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роуз, Дж. К., Браам, Дж. , Фрай, С. К., и Нишитани, К. (2002). Семейство ферментов XTH, участвующих в эндотрансглюкозилировании и эндогидролизе ксилоглюкана: современные перспективы и новая объединяющая номенклатура. Физиол клеток растений. 43, 1421–1435. doi: 10.1093/pcp/pcf171
, Фрай, С. К., и Нишитани, К. (2002). Семейство ферментов XTH, участвующих в эндотрансглюкозилировании и эндогидролизе ксилоглюкана: современные перспективы и новая объединяющая номенклатура. Физиол клеток растений. 43, 1421–1435. doi: 10.1093/pcp/pcf171
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сантос, А.П., Серра Т., Фигейреду Д. Д., Баррос П., Лоренсу Т., Чандер С. и др. (2011). Транскрипционная регуляция реакции на абиотический стресс у риса: комбинированное действие факторов транскрипции и эпигенетических механизмов. ОМИКС 15, 839–857. doi: 10.1089/omi.2011.0095
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Секи М., Нарусака М., Исида Дж., Нанджо Т., Фудзита М., Ооно Ю. и др. (2002). Мониторинг профилей экспрессии 7000 генов арабидопсиса в условиях засухи, холода и сильного засоления с использованием полноразмерного микрочипа кДНК. Завод J. 31, 279–292. doi: 10.1046/j. 1365-313X.2002.01359.x
1365-313X.2002.01359.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стоун, Б.А., и Кларк, А.Е. (1992). Химия и биология (1 → 3)-β-D-глюканов . Мельбурн, Виктория: Издательство Университета Ла Троб.
Академия Google
Svensson, J.T., Crosatti, C., Campoli, C., Bassi, R., Stanca, A.M., Close, T.J., et al. (2006). Транскриптомный анализ адаптации к холоду у мутантов ячменя Albina и Xantha. Завод физиол. 141, 257–270. doi: 10.1104/стр.105.072645
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Суонстон, Дж. С., Эллис, Р. П., Перес-Вендрель, А., Вольтас, Дж., и Молина-Кано, Дж. Л. (1997). Закономерности развития зерна ячменя в Испании и Шотландии и их влияние на качество солода. Зерновые хим. 74, 456–461. doi: 10.1094/CCHEM.1997.74.4.456
Полнотекстовая перекрестная ссылка | Академия Google
Такета, С., Юо Т., Тонука Т., Цумурая Ю., Инагаки Ю., Харуяма Н. и др. (2012). Функциональная характеристика бетаглюкановых мутантов ячменя демонстрирует уникальную роль CslF6 в биосинтезе (1, 3; 1, 4)-β-D-глюкана. Дж. Экспл. Бот. 63, 381–392. дои: 10.1093/jxb/err285
и др. (2012). Функциональная характеристика бетаглюкановых мутантов ячменя демонстрирует уникальную роль CslF6 в биосинтезе (1, 3; 1, 4)-β-D-глюкана. Дж. Экспл. Бот. 63, 381–392. дои: 10.1093/jxb/err285
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Taylor-Teeples, M., Lin, L., de Lucas, M., Turco, G., Toal, T.W., Gaudinier, A., et al. (2015). Регуляторная сеть генов арабидопсиса для синтеза вторичной клеточной стенки. Природа 517, 571–575. doi: 10.1038/nature14099
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Такер, М. Р., и Колтунов, А. М. (2014). Мониторы трафика на периферии клетки: роль клеточных стенок во время ранней дифференцировки женских половых клеток у растений. Курс. мнение биол. растений 17, 137–145. doi: 10.1016/j.pbi.2013.11.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тернер, С. Р., и Сомервилл, К.Р. (1997). Фенотип коллапсированной ксилемы арабидопсиса идентифицирует мутантов с дефицитом отложения целлюлозы во вторичной клеточной стенке. Растительная клетка 9, 689–701. doi: 10.1105/tpc.9.5.689
Фенотип коллапсированной ксилемы арабидопсиса идентифицирует мутантов с дефицитом отложения целлюлозы во вторичной клеточной стенке. Растительная клетка 9, 689–701. doi: 10.1105/tpc.9.5.689
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Турра, Д., Эль Галид, М., Росси, Ф., и Ди Пьетро, А. (2015). Грибной патоген использует рецептор полового феромона для хемотропного восприятия сигналов растения-хозяина. Природа 527, 521–524. doi: 10.1038/nature15516
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ван Дам, Н.М. (2009). Подземные травоядные и защита растений. год. Преподобный Экол. Эвол. Сист. 40, 373–391. doi: 10.1146/annurev.ecolsys.110308.120314
Полнотекстовая перекрестная ссылка | Академия Google
Ванденборре, Г., Смагге, Г., и Ван Дамм, Э. Дж. (2011). Лектины растений как защитные белки от насекомых-фитофагов. Фитохимия 72, 1538–1550. doi: 10.1016/j. phytochem.2011.02.024
phytochem.2011.02.024
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вега-Санчес, М.E., Verhertbruggen, Y., Christensen, U., Chen, X., Sharma, V., Varanasi, P., et al. (2012). Потеря целлюлозосинтазоподобной функции F6 влияет на отложение глюкана со смешанной связью, механические свойства клеточной стенки и защитные реакции в вегетативных тканях риса. Завод физиол. 159, 56–69. doi: 10.1104/стр.112.195495
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Vogel, J.P., Raab, T.K., Somerville, C.R., and Somerville, S.C. (2002). PMR6 , ген, подобный пектатлиазе, необходимый для восприимчивости арабидопсиса к мучнистой росе. Растительная клетка 14, 2095–2106. doi: 10.1105/tpc.003509
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Vogel, JP, Raab, T.K., Somerville, CR, and Somerville, SC (2004). Мутации в PMR5 приводят к устойчивости к мучнистой росе и изменению состава клеточных стенок. Завод J. 40, 968–978. doi: 10.1111/j.1365-313X.2004.02264.х
Завод J. 40, 968–978. doi: 10.1111/j.1365-313X.2004.02264.х
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Уоллворк, М.А.Б., Дженнер, К.Ф., Лог, С.Дж., и Седжли, М. (1998). Влияние высокой температуры при наливе зерна на структуру формирующегося и соложеного зерна ячменя. Энн. Бот. 82, 587–599. doi: 10.1006/anbo.1998.0721
Полнотекстовая перекрестная ссылка | Академия Google
Ван, Т., Макфарлейн, Х. Э., и Перссон, С. (2016). Влияние абиотических факторов на синтез целлюлозы. Дж.Эксп. Бот. 67, 543–552. дои: 10.1093/jxb/erv488
Реферат PubMed | Полнотекстовая перекрестная ссылка
Wang, Y., Yang, L., Zheng, Z., Grumet, R., Loescher, W., Zhu, J.K., et al. (2013). Транскриптомные и физиологические вариации трех экотипов арабидопсиса в ответ на солевой стресс. PLoS ONE 8:e69036. doi: 10.1371/journal.pone.0069036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вечорек, К. (2015). Глава Трехклеточная стенка пораженных нематодами корней. Доп. Бот. Рез. 73, 61–90. doi: 10.1016/bs.abr.2014.12.002
(2015). Глава Трехклеточная стенка пораженных нематодами корней. Доп. Бот. Рез. 73, 61–90. doi: 10.1016/bs.abr.2014.12.002
Полнотекстовая перекрестная ссылка | Академия Google
Wieczorek, K., Elashry, A., Quentin, M., Grundler, F.M.W., Favery, B., Seifert, G.J., et al. (2014). Определена роль пектатлиаз в формировании питающих структур, индуцируемых цистными и галловыми нематодами. Мол. Взаимодействие растительных микробов. 27, 901–912. doi: 10.1094/MPMI-01-14-0005-R
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ву, М.О., Бирд Х., Макдональд М.Х., Брюэр Э.П., Юссеф Р.М., Ким Х. и соавт. (2014). Манипуляции с двумя генами α-эндо-β-1,4-глюканазы, AtCel6 и GmCel7, снижают восприимчивость корней сои к глицинам Heterodera. Мол. Завод Патол. 15, 927–939. doi: 10.1111/mpp.12157
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ву, Ю., и Косгроув, Д. Дж. (2000). Адаптация корней к низкому водному потенциалу за счет изменения растяжимости клеточной стенки и белков клеточной стенки. Дж. Экспл. Бот. 51, 1543–1553. doi: 10.1093/jexbot/51.350.1543
Дж. Экспл. Бот. 51, 1543–1553. doi: 10.1093/jexbot/51.350.1543
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Инь, Ю. Б., Монен, Д., Гелинео-Альбершайм, И., Сюй, Ю. и Хан, М. Г. (2011). Гликозилтрансферазы семейства GT8. год. Заводской номер . 41, 167–211. дои: 10.1002/97814443
.ch6
Полнотекстовая перекрестная ссылка | Академия Google
Чжэн, Дж., Фу, Дж., Гоу, М., Хуай, Дж., Лю, Ю., Цзянь, М., и др. (2010). Полногеномный транскриптомный анализ двух инбредных линий кукурузы в условиях засушливого стресса. Завод Мол. биол. 72, 407–421. doi: 10.1007/s11103-009-9579-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжун, Р., Кейс, С.Дж., Шредер, Б.П., и Йе, З.Х. (2002). Мутация хитиназоподобного гена вызывает эктопическое отложение лигнина, аберрантную форму клеток и перепроизводство этилена. Растительная клетка 14, 165–179. doi: 10.1105/tpc.010278
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Синтетическая 5,3-сшивка в клеточной стенке палочковидных грамположительных бактерий
Грамположительные бактерии являются монодермами, и их единственная мембрана окружена многослойной (15–30 нм) клеточной стенкой. Напротив, грамотрицательные бактерии являются дидермами, а их клеточная стенка одно- или двухслойная (3–6 нм) и располагается между внутренней и внешней мембранами (1). Несмотря на это несоответствие в общей архитектуре, химические структуры пептидогликана каждого организма удивительно похожи (2). Пептидогликан клеточной стенки, или муреин, состоит из гликановых нитей, собранных из сахаридных составляющих N -ацетилмурамовой кислоты (NAM) и N -ацетилглюкозамина (NAG), и в каждом классе бактерий пентапептид простирается от лактильная группа сахарида НАМ (рис.1 А ) (3). В то время как вариабельность последовательности пептидного стебля существует у разных видов, архетипическая структура стебля монодермы представляет собой l-Ala-γ-d-Glu-l-Lys-d-Ala-d-Ala, а в дидерме l-Lys обычно замещен карбоксипроизводным мезо -2,6-диаминопимелат ( m -DAP) (рис. 1 B ) (3, 4). Пептидные стержни клеточной стенки поперечно связаны пенициллин-связывающим белком (PBP) d,d-транспептидазами и l,d-транспептидазами.
Напротив, грамотрицательные бактерии являются дидермами, а их клеточная стенка одно- или двухслойная (3–6 нм) и располагается между внутренней и внешней мембранами (1). Несмотря на это несоответствие в общей архитектуре, химические структуры пептидогликана каждого организма удивительно похожи (2). Пептидогликан клеточной стенки, или муреин, состоит из гликановых нитей, собранных из сахаридных составляющих N -ацетилмурамовой кислоты (NAM) и N -ацетилглюкозамина (NAG), и в каждом классе бактерий пентапептид простирается от лактильная группа сахарида НАМ (рис.1 А ) (3). В то время как вариабельность последовательности пептидного стебля существует у разных видов, архетипическая структура стебля монодермы представляет собой l-Ala-γ-d-Glu-l-Lys-d-Ala-d-Ala, а в дидерме l-Lys обычно замещен карбоксипроизводным мезо -2,6-диаминопимелат ( m -DAP) (рис. 1 B ) (3, 4). Пептидные стержни клеточной стенки поперечно связаны пенициллин-связывающим белком (PBP) d,d-транспептидазами и l,d-транспептидазами. У грамотрицательных бактерий эпсилон-аминогруппа m -DAP служит нуклеофилом, замещающим концевой d-Ala соседнего пептидного стебля при образовании катализируемых d,d-транспептидазой 4,3- и l,d катализируемые транспептидазой 3,3-пептидные поперечные связи (5).Обе структуры поперечного связывания стабилизируют клеточную стенку, а перекрестная связь l,d-транспептидазы дополнительно закрепляет клеточную стенку на внешней мембране через липопротеин Брауна (6). У грамположительных бактерий эпсилон-аминогруппа лизина действует как соответствующий нуклеофил при образовании поперечных связей. Из-за отсутствия внешней мембраны у грамположительных бактерий поперечное связывание l,d-транспептидазой является редким событием, которое может помочь в обеспечении механической стабильности. Функциональная избыточность очевидна в синтезе клеточной стенки, поскольку Escherichia coli кодирует не менее пяти d,d-транспептидаз и шесть l,d-транспептидаз, а Bacillus subtilis кодирует не менее 10 d,d-транспептидаз и две l,d-транспептидазы.
У грамотрицательных бактерий эпсилон-аминогруппа m -DAP служит нуклеофилом, замещающим концевой d-Ala соседнего пептидного стебля при образовании катализируемых d,d-транспептидазой 4,3- и l,d катализируемые транспептидазой 3,3-пептидные поперечные связи (5).Обе структуры поперечного связывания стабилизируют клеточную стенку, а перекрестная связь l,d-транспептидазы дополнительно закрепляет клеточную стенку на внешней мембране через липопротеин Брауна (6). У грамположительных бактерий эпсилон-аминогруппа лизина действует как соответствующий нуклеофил при образовании поперечных связей. Из-за отсутствия внешней мембраны у грамположительных бактерий поперечное связывание l,d-транспептидазой является редким событием, которое может помочь в обеспечении механической стабильности. Функциональная избыточность очевидна в синтезе клеточной стенки, поскольку Escherichia coli кодирует не менее пяти d,d-транспептидаз и шесть l,d-транспептидаз, а Bacillus subtilis кодирует не менее 10 d,d-транспептидаз и две l,d-транспептидазы. -транспептидазы (5, 7, 8).Эта избыточность подразумевает, что бактерии развили тщательный баланс в сборке клеточной стенки, который образует идеальный биополимер для бактериальной жизни.
-транспептидазы (5, 7, 8).Эта избыточность подразумевает, что бактерии развили тщательный баланс в сборке клеточной стенки, который образует идеальный биополимер для бактериальной жизни.
Расположение пептидных поперечных связей определяет размеры, пропорции и пористость клеточной стенки, а также форму бактерии. Чтобы изучить структурные ограничения архитектуры клеточной стенки, которые позволяют ей тщательно управлять основными биологическими процессами и поддерживать клеточную морфологию, мы начали заменять естественные канонические перекрестные связи клеточной стенки (рис.1 B ) с неестественными синтетическими поперечными связями клеточной стенки. Ранее мы использовали электрофильные неканонические d-аминокислоты (d-AA) для образования неестественных синтетических 4,3-сшивок клеточной стенки в грамотрицательной бактерии E. coli (9). Здесь мы распространяем этот подход на грамположительные бактерии B. subtilis , выявляя различия в расположении синтетических поперечных связей, индуцированных синтетическими поперечными связями эффектах на клеточный рост и фенотип, а также различия в способе накопления синтетическая клеточная стенка бактерий.
Результаты и обсуждение
Экспериментальный подход.
Наши предыдущие исследования образования неестественных поперечных связей клеточной стенки у грамотрицательных бактерий зависели от способности бактериальных транспептидаз включать d-АК из окружающей среды в клеточную стенку, стратегия, используемая бактериями для уменьшения количества пептидов. сшивание (рис. 2 A ) (9⇓⇓–12). В оригинальных исследованиях другие использовали этот подход для включения флуорофоров и биортогональных фото-сшивающих агентов в клеточную стенку различных бактерий для визуализации клеточной стенки живых клеток (13, 14).Используя неканонические электрофильные d-AA в качестве субстратов для транспептидаз, мы смогли заменить ~30% естественных поперечных связей клеточной стенки в E. coli неестественными синтетическими поперечными связями клеточной стенки без сопутствующего наблюдаемого эффекта. на бактериальный фенотип (9). Чтобы определить, можно ли использовать подобную стратегию для образования неестественных перекрестных связей клеточной стенки у грамположительных бактерий, мы выбрали B. subtilis . B. subtilis является модельной бактерией для изучения структуры палочковидных спорообразующих грамположительных бактерий (15).Ключевым отличием клеточной стенки B. subtilis от других грамположительных бактерий является присутствие архетипического грамотрицательного m -DAP в третьем положении пептидного стебля, который у B. subtilis обычно существует в виде катализируемой ферментами амидированной аминокислоты (16).
subtilis . B. subtilis является модельной бактерией для изучения структуры палочковидных спорообразующих грамположительных бактерий (15).Ключевым отличием клеточной стенки B. subtilis от других грамположительных бактерий является присутствие архетипического грамотрицательного m -DAP в третьем положении пептидного стебля, который у B. subtilis обычно существует в виде катализируемой ферментами амидированной аминокислоты (16).
( A ) Сравнительные механизмы опосредованного d,d-транспептидазой канонического перекрестного связывания клеточной стенки и неканонического перекрестного связывания клеточной стенки экзогенными d-AA в B.субтилис . ( B ) Структуры электрофильных неканонических d-AA и родственных контролей l-AA. ( C ) Кривые роста бактерий B. subtilis , необработанных или обработанных d-AA 1c или l-AA 1d . ( D ) Структура неканонического 5,3-сшитого NAG-NAM-(пентапептид)-NAG-ангидроNAM-(тетрапептид), образованного 1c . Серые прямоугольники показывают сравнение синтетической сшивки и нативной сшивки. Для каждого неканонического d-AA образующийся первичный синтетический сшитый муропептид включает канонический NAG-NAM-(тетрапептид) (R’) и NAG-ангидроNAM-(тетрапептид) (R»), где установлен неканонический d-AA. рядом с d-Ala в четвертой позиции стержня R ‘, заменяя d-Ala в пятой позиции.Масс-спектры, соответствующие синтетическим несшитым и сшитым видам клеточной стенки, образованным ( E , F ) 1a , ( G , H ) 1b , и ( I , J ) 1c показаны. Соединения 2a и 2b образуют синтетические несшитые муропептиды, но не синтетические сшитые муропептиды. Структура и массы каждого неестественного синтетического муропептида клеточной стенки представлены в приложении SI (приложение SI , рис.С3–С8).
Серые прямоугольники показывают сравнение синтетической сшивки и нативной сшивки. Для каждого неканонического d-AA образующийся первичный синтетический сшитый муропептид включает канонический NAG-NAM-(тетрапептид) (R’) и NAG-ангидроNAM-(тетрапептид) (R»), где установлен неканонический d-AA. рядом с d-Ala в четвертой позиции стержня R ‘, заменяя d-Ala в пятой позиции.Масс-спектры, соответствующие синтетическим несшитым и сшитым видам клеточной стенки, образованным ( E , F ) 1a , ( G , H ) 1b , и ( I , J ) 1c показаны. Соединения 2a и 2b образуют синтетические несшитые муропептиды, но не синтетические сшитые муропептиды. Структура и массы каждого неестественного синтетического муропептида клеточной стенки представлены в приложении SI (приложение SI , рис.С3–С8).
У грамотрицательных бактерий неканонические d-AA включаются в пептидный стебель в четвертом положении с помощью l,d-транспептидаз и в пятом положении с помощью d,d-транспептидаз (17). В наших предыдущих усилиях по созданию неестественных синтетических перекрестных связей клеточной стенки мы в первую очередь наблюдали замену аминокислот в четвертом положении, вероятно, в результате удаления d,d-карбоксипептидазами любых неканонических d-AA, включенных в пятое положение стеблевого пептида. (9, 17). Однако у грамположительных бактерий l,d-транспептидазы редко образуют поперечные связи, что ограничивает возможность включения неканонических d-AA в четвертое положение (17, 18).
В наших предыдущих усилиях по созданию неестественных синтетических перекрестных связей клеточной стенки мы в первую очередь наблюдали замену аминокислот в четвертом положении, вероятно, в результате удаления d,d-карбоксипептидазами любых неканонических d-AA, включенных в пятое положение стеблевого пептида. (9, 17). Однако у грамположительных бактерий l,d-транспептидазы редко образуют поперечные связи, что ограничивает возможность включения неканонических d-AA в четвертое положение (17, 18).
В качестве альтернативного подхода ранее было показано, что нокаут гена dacA , который кодирует первичную d,d-карбоксипептидазу PBP5, позволяет накапливать флуоресцентные d-AA в пятой позиции пептидных стеблей B. subtilis (13). Мы пришли к выводу, что та же самая стратегия может быть использована для включения неканонических электрофильных d-AA в пятое положение пептидных стеблей B. subtilis Δ dacA . Эти d-AA могут быть расположены соответствующим образом для ковалентной перекрестной связи с амидированным m -DAP соседнего пептидного стебля, что приводит к неестественным синтетическим 5,3-поперечным связям (рис. 2 А ). С этой целью мы синтезировали пять d-АК в качестве потенциальных структур для образования синтетических поперечных связей (9). Аминокислоты 1a , 1b и 1c являются фторсульфатами, способными реагировать с амино- и гидроксильными группами аминокислот в непосредственной близости, а 2a и 2b являются несколько более реакционноспособными винилсульфонамидами (рис. 2 9009) В ) (19). Варианты с различными паттернами замещения колец и длинами боковых цепей оценивали на эффективность включения в клеточные стенки.Ранее мы показали, что длина боковой цепи и природа электрофила значительно влияют на включение d-AA и эффективность образования синтетических поперечных связей. Кроме того, в качестве контроля были приготовлены l-энантиомеры 1c ( 1d ) и 1a ( 1e ).
2 А ). С этой целью мы синтезировали пять d-АК в качестве потенциальных структур для образования синтетических поперечных связей (9). Аминокислоты 1a , 1b и 1c являются фторсульфатами, способными реагировать с амино- и гидроксильными группами аминокислот в непосредственной близости, а 2a и 2b являются несколько более реакционноспособными винилсульфонамидами (рис. 2 9009) В ) (19). Варианты с различными паттернами замещения колец и длинами боковых цепей оценивали на эффективность включения в клеточные стенки.Ранее мы показали, что длина боковой цепи и природа электрофила значительно влияют на включение d-AA и эффективность образования синтетических поперечных связей. Кроме того, в качестве контроля были приготовлены l-энантиомеры 1c ( 1d ) и 1a ( 1e ).
Формирование синтетических поперечных связей неканоническими d-AA.

Сначала мы определили, являются ли неканонические d-AA токсичными для B. subtilis Δ dacA при повышенных концентрациях.L-энантиомеры фторсульфата и винилсульфонамида, замещенного фенилаланина, ранее были включены в белки живых бактерий с минимальным влиянием на рост бактерий при концентрации 1 мМ (19, 20). В E. coli добавление d-АК и родственного им L-энантиомера в среду для выращивания приводило к идентичным кривым роста со значительным влиянием на скорость роста, начиная с концентраций выше 8 мМ (9). Однако у B. subtilis Δ dacA мы наблюдали повышенное вредное воздействие на рост клеток неканонических d-AA по сравнению с l-AA, начиная с 4 мМ (рис.2 C и SI Приложение , рис. S1 A – H ). Соединения 1A , 1B , 1C , 1D , 1E , 2A и 2B полностью ингибируют рост на 8 мм, 8 мм, 8 мм,> 8 мм,> 8 мм, Концентрации >8 мМ, >8 мМ соответственно.
Чтобы определить, включены ли неканонические d-AA в клеточную стенку, мы сначала обработали культуры B. subtilis Δ dacA 1 мМ каждого d-AA.Вкратце, ночную культуру B. subtilis Δ dacA инокулировали в среду Luria Bertani (LB) с добавлением хлорамфеникола и выращивали до оптической плотности 600 нм (OD 600 ) = 0,05. Затем культуры независимо обрабатывали 1 мМ каждого d-AA и выращивали до OD 600 = 1,10. Затем клетки помещали на лед, осаждали и выделяли клеточную стенку каждой культуры в соответствии с модифицированной версией установленной методики (9, 21, 22).Мы проанализировали каждый образец с помощью жидкостной хроматографии и масс-спектрометрии (ЖХ-МС). Концентрация каждого образца была нормализована к наиболее распространенному нативному несшитому муропептиду, NAG-NAM-пентапептиду, при сигнале масс-спектра ~10 6 ( SI Приложение , рис. S2 A и B ). Массы, соответствующие включению неканонических d-АК в пятое положение пептидных стеблей, были обнаружены для каждого соединения и предоставлен соответствующий сигнал масс-спектров: 1a (10 6 ), 1b (10 6 ), 1c (10 6 ), 2a (10 5 ) и 2b (10 6 ) (рис. 2 E , 2 G , и 2 I , и SI Приложение , рис. S2 C и D ). В образцах, обработанных контрольными I-AA 1d и 1e , включения не обнаружено.
2 E , 2 G , и 2 I , и SI Приложение , рис. S2 C и D ). В образцах, обработанных контрольными I-AA 1d и 1e , включения не обнаружено.
Затем мы проанализировали данные МС для массы, соответствующей синтетической поперечной сшивке. Ожидаемая структура синтетической 5,3-сшивки, образованной 1c , показана на рис. 2 D . Масс-спектральный анализ подтвердил ожидаемые синтетические 5,3-сшивки клеточной стенки для всех трех фторсульфатов 1a , 1b и 1c (полный список структур приведен в SI, Приложение , рис.С3–С8). Структура синтетической 5,3-сшивки, образованной 1c , была подтверждена методом ЖХ-МС/МС ( SI Приложение , рис. S9). Синтетические сшитые муропептиды, образованные 1a , 1b и 1c , давали сравнимые сигналы масс-спектров ~10 6 и были примерно в 4 раза ниже, чем сигнал, наблюдаемый для синтетических несшитых пептидов. муропептиды (рис. 2 E – J ). Синтетический сшитый муропептид не обнаружен для винилсульфонамидов 2a или 2b , которые ранее образовывали синтетические сшивки в E.coli (9). Кроме того, мы не обнаруживаем продукта реакции m -DAP и свободной d-AA.
муропептиды (рис. 2 E – J ). Синтетический сшитый муропептид не обнаружен для винилсульфонамидов 2a или 2b , которые ранее образовывали синтетические сшивки в E.coli (9). Кроме того, мы не обнаруживаем продукта реакции m -DAP и свободной d-AA.
Примечательно, что в наших предыдущих исследованиях с E. coli 1c плохо включался и гораздо менее эффективно образовывал поперечные связи по сравнению с 1a или 1b (9). Это несоответствие в эффективности сшивания, вероятно, связано с пространственным расположением пептидных стеблей, таким образом, удлиненная боковая цепь 1c помещает фторсульфат в непосредственной близости к эпсилон-аминогруппе m -DAP в B.субтилис . Поскольку неканонические d-AA включены в четвертую позицию в E. coli , это удлинение, вероятно, ухудшает взаимодействие с m -DAP соседнего пептидного стебля. Неожиданной особенностью нашего анализа является то, что донорная цепь м -DAP первичного сшитого муропептида d-AA представляет собой тетрапептид (рис. 2 D ), в то время как в первичном нативном сшитом муропептиде эта же цепь представляет собой пентапептид ( SI Приложение , рис. S3).В то время как клетка кодирует другие низкоактивные d,d-карбоксипептидазы (23), это структурное несоответствие было неожиданным для штамма Δ dacA и может указывать на ключевые различия в процессинге клеточной стенки для двух муропептидов. Затем мы сосредоточились на дальнейшей характеристике поперечных связей, образованных d-AA 1c .
Неожиданной особенностью нашего анализа является то, что донорная цепь м -DAP первичного сшитого муропептида d-AA представляет собой тетрапептид (рис. 2 D ), в то время как в первичном нативном сшитом муропептиде эта же цепь представляет собой пентапептид ( SI Приложение , рис. S3).В то время как клетка кодирует другие низкоактивные d,d-карбоксипептидазы (23), это структурное несоответствие было неожиданным для штамма Δ dacA и может указывать на ключевые различия в процессинге клеточной стенки для двух муропептидов. Затем мы сосредоточились на дальнейшей характеристике поперечных связей, образованных d-AA 1c .
Количественное определение состава синтетической клеточной стенки.
Мы оценили влияние различных концентраций 1c на плотность синтетических поперечных связей клеточной стенки.Культуры готовили, как описано ранее, и независимо обрабатывали 1, 2 и 4 мМ 1c . Дополнительно мы подготовили необработанную культуру и культуру, обработанную соединением 1d , в качестве контроля. После экстракции и переваривания клеточных стенок образцы анализировали с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). При самой высокой испытанной концентрации (4 мМ 1c ) процент наиболее распространенного неканонического d-AA, содержащего синтетические несшитые виды (SnC), амидированного NAG-NAM-тетрапептида- 1c , составлял ~19% от общего несшитого пептидогликана; наиболее распространенными нативными несшитыми видами (nC) были амидированные NAG-NAM-пентапептиды (рис.3 A и B ). Первичная d-АК, содержащая синтетические сшитые частицы (СК), амидированный NAG-NAM-тетрапептид- 1c -тетрапептид-NAG-ангидроNAM (структура на рис. 2 D ), составляла 19% от общего количества сшитый пептидогликан; наиболее распространенными природными сшитыми видами (C) были амидированные NAG-NAM-тетрапептид-пентапептид-NAG-NAM (фиг. 3 A и B ). Анализ ВЭЖХ показал зависящее от концентрации увеличение количества d-AA-содержащих видов клеточной стенки SnC и SC (фиг.
После экстракции и переваривания клеточных стенок образцы анализировали с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ). При самой высокой испытанной концентрации (4 мМ 1c ) процент наиболее распространенного неканонического d-AA, содержащего синтетические несшитые виды (SnC), амидированного NAG-NAM-тетрапептида- 1c , составлял ~19% от общего несшитого пептидогликана; наиболее распространенными нативными несшитыми видами (nC) были амидированные NAG-NAM-пентапептиды (рис.3 A и B ). Первичная d-АК, содержащая синтетические сшитые частицы (СК), амидированный NAG-NAM-тетрапептид- 1c -тетрапептид-NAG-ангидроNAM (структура на рис. 2 D ), составляла 19% от общего количества сшитый пептидогликан; наиболее распространенными природными сшитыми видами (C) были амидированные NAG-NAM-тетрапептид-пентапептид-NAG-NAM (фиг. 3 A и B ). Анализ ВЭЖХ показал зависящее от концентрации увеличение количества d-AA-содержащих видов клеточной стенки SnC и SC (фиг. 3 B и SI Приложение , рис. С10–С12). Эти результаты показывают, что B. subtilis выживает в условиях (4 мМ 1c ), где 19% клеточной стенки заменены неканоническими строительными блоками. При 2 мМ соединения 1c , B . subtilis растет спокойно, а клеточная стенка содержит 12% синтетических поперечных связей.
3 B и SI Приложение , рис. С10–С12). Эти результаты показывают, что B. subtilis выживает в условиях (4 мМ 1c ), где 19% клеточной стенки заменены неканоническими строительными блоками. При 2 мМ соединения 1c , B . subtilis растет спокойно, а клеточная стенка содержит 12% синтетических поперечных связей.
( A ) ВЭЖХ-кривая (205 нм) выделенного пептидогликана из B. subtilis , обработанного 1c (4 мМ).Помечены пики, соответствующие наиболее распространенным видам муропептидов для нативного несшитого (nC), нативного сшитого (C), синтетического несшитого (SnC) и синтетического сшитого (SC). SnC и SC соответствуют структурам, модифицированным d-AA. Черные кружки обозначают амидированные виды, серые кружки обозначают неамидированные несшитые виды, а золотые кружки обозначают частично амидированные и неамидированные сшитые виды. ( B ) Процент синтетических несшитых и синтетических сшитых муропептидов для клеточной стенки, выделенных из B. subtilis после обработки 1c . Обратите внимание, проценты основаны исключительно на наиболее распространенных амидированных местных и синтетических видах. Подробная количественная оценка представлена в SI Приложение .
subtilis после обработки 1c . Обратите внимание, проценты основаны исключительно на наиболее распространенных амидированных местных и синтетических видах. Подробная количественная оценка представлена в SI Приложение .
Бактериальный фенотип, полученный путем перекрестной связи d-AA.
Мы проанализировали B. subtilis с помощью сканирующей электронной микроскопии (СЭМ) на предмет наблюдаемого изменения клеточной морфологии в результате неестественных синтетических поперечных связей. B. subtilis Δ dacA выращивали до OD 600 = 0.05 и обрабатывали 1 мМ, 2 мМ или 4 мМ 1c или 1d в течение 2 ч; необработанные клетки дикого типа и Δ dacA использовали в качестве контроля. Клетки были подготовлены для SEM-анализа, как описано ранее (9). Клетки дикого типа B. subtilis имеют характерную линейную стержневую форму с делением в середине клетки (рис. 4 A ). Для сравнения, B. subtilis Δ dacA немного короче, но в остальном имеет сходную морфологию (рис. 4 B ).Ранее сообщалось о наблюдении, что B. subtilis Δ dacA демонстрирует укороченную морфологию, и литература предполагает, что укорочение клеток становится более глубоким, когда клетки входят в стационарную фазу (24). Напротив, B. subtilis Δ dacA , обработанные 4 мМ 1c , подвергаются значительному лизису клеток, что частично показано белыми стрелками (фиг. 4 C и E ). Клетки, которые выживают после обработки, выглядят удлиненными и спиралевидными (рис.4 C – E ). Для сравнения, клетки, обработанные l-энантиомером 1d , успешно делятся и имеют длину, сравнимую с необработанными клетками (рис. 4. к нецелевому взаимодействию фторсульфата с клеточными компонентами. Этот эффект скручивания в сочетании с удлинением клеток, вызванным обработкой 1c , может способствовать наблюдаемому спиралевидному вращению (рис.
4 A ). Для сравнения, B. subtilis Δ dacA немного короче, но в остальном имеет сходную морфологию (рис. 4 B ).Ранее сообщалось о наблюдении, что B. subtilis Δ dacA демонстрирует укороченную морфологию, и литература предполагает, что укорочение клеток становится более глубоким, когда клетки входят в стационарную фазу (24). Напротив, B. subtilis Δ dacA , обработанные 4 мМ 1c , подвергаются значительному лизису клеток, что частично показано белыми стрелками (фиг. 4 C и E ). Клетки, которые выживают после обработки, выглядят удлиненными и спиралевидными (рис.4 C – E ). Для сравнения, клетки, обработанные l-энантиомером 1d , успешно делятся и имеют длину, сравнимую с необработанными клетками (рис. 4. к нецелевому взаимодействию фторсульфата с клеточными компонентами. Этот эффект скручивания в сочетании с удлинением клеток, вызванным обработкой 1c , может способствовать наблюдаемому спиралевидному вращению (рис. 4 D и E ). B. subtilis Δ dacA , обработанные 1 мМ 1c или 1d , не обнаруживают значительных изменений в морфологии, в то время как клетки, обработанные 2 мМ 1c или 1d , демонстрируют небольшое скручивание, которое демонстрирует небольшое скручивание, обработка 1c . Дополнительные изображения SEM для каждой концентрации соединения приведены в SI, Приложение , рис. S13 A – F . В совокупности эти данные показывают, что клетки, содержащие синтетические поперечные связи клеточной стенки, удлиняются и, вероятно, имеют нарушенную способность к делению.
4 D и E ). B. subtilis Δ dacA , обработанные 1 мМ 1c или 1d , не обнаруживают значительных изменений в морфологии, в то время как клетки, обработанные 2 мМ 1c или 1d , демонстрируют небольшое скручивание, которое демонстрирует небольшое скручивание, обработка 1c . Дополнительные изображения SEM для каждой концентрации соединения приведены в SI, Приложение , рис. S13 A – F . В совокупности эти данные показывают, что клетки, содержащие синтетические поперечные связи клеточной стенки, удлиняются и, вероятно, имеют нарушенную способность к делению.
B. subtilis культивировали до OD 600 = 0,05 и либо не обрабатывали, либо обрабатывали 4 мМ 1c или 1d в течение 2 часов. Бактерии были визуализированы с помощью SEM, и результаты показаны. СЭМ необработанных бактерий ( A ) B. subtilis дикого типа и ( B ) B. subtilis ΔdacA демонстрирует линейную стержневую форму. Мутация ΔdacA дает слегка укороченные клетки. Изображения B. subtilis ΔdacA , обработанного 1c , были получены при увеличении ( C ) 1500× и ( D , E ) 15000×.Наблюдается значительный лизис клеток, как указано белыми стрелками. Бактерии, которые выживают после обработки 1c , способны успешно удлиняться, но деление клеток оказывается нарушенным. Удлиненные клетки имеют спиральный фенотип. Изображения B. subtilis ΔdacA , обработанного 1d , были получены при увеличении ( F ) 1500× и ( G , H ) 15000×. Для бактерий, обработанных 1d , лизиса клеток не наблюдалось, хотя клетки проявляли едва заметный изгиб.Белая полоса масштаба 1 мкМ показана в A , B , D , E , G и H ( вверху справа ). Белая шкала 5 мкМ показана в C и F ( вверху справа ).
subtilis дикого типа и ( B ) B. subtilis ΔdacA демонстрирует линейную стержневую форму. Мутация ΔdacA дает слегка укороченные клетки. Изображения B. subtilis ΔdacA , обработанного 1c , были получены при увеличении ( C ) 1500× и ( D , E ) 15000×.Наблюдается значительный лизис клеток, как указано белыми стрелками. Бактерии, которые выживают после обработки 1c , способны успешно удлиняться, но деление клеток оказывается нарушенным. Удлиненные клетки имеют спиральный фенотип. Изображения B. subtilis ΔdacA , обработанного 1d , были получены при увеличении ( F ) 1500× и ( G , H ) 15000×. Для бактерий, обработанных 1d , лизиса клеток не наблюдалось, хотя клетки проявляли едва заметный изгиб.Белая полоса масштаба 1 мкМ показана в A , B , D , E , G и H ( вверху справа ). Белая шкала 5 мкМ показана в C и F ( вверху справа ).
Документально подтверждено, что пептидогликан в перегородке во время клеточного деления часто лишается пептидных стеблей, которые клипируются периплазматическими амидазами по лактильной группе (25). Сахариды в перегородке направляются и ремоделируются белками клеточного деления дивисомы, а также некоторыми ферментами ремоделирования клеточной стенки, которые содержат домены повторов, связанных со споруляцией (SPOR), которые распознают и связываются с обнаженным гликаном (25⇓–27). ).Кроме того, делеция гена амидаз периплазматической клеточной стенки приводит к фенотипу удлиненных клеток (28). В нашем анализе ЖХ-МС мы наблюдаем продукт реакции первичной нативной поперечной связи с амидазами клеточной стенки, но не продукт реакции первичной синтетической поперечной связи с амидазами клеточной стенки на обоих концах (т.е. NAG- НАМ или НАГ-ангидроНАМ). Мы предполагаем, что присутствие синтетически модифицированной клеточной стенки препятствует активности периплазматических амидаз и, следовательно, ферментов, содержащих домен SPOR, связанных с клеточным делением, что приводит к нарушению клеточного деления и наблюдаемому фенотипу при повышенных концентрациях 1c .В настоящее время мы не знаем, почему это наблюдение уникально для нашей модели грамположительной бактерии B. subtilis и не обнаружено в нашей модели грамотрицательной бактерии E . coli , но предполагают, что это может быть связано со специфичностью субстрата и объемом необходимых ферментов, участвующих в делении.
Интригует фенотипическое сравнение спиралевидности наших клеток B. subtilis с таковой у Campylobacter . Спиралевидная форма Campylobacter контролируется белками, определяющими форму клетки (Csd), которые часто содержат домены лизостафинподобных металлопротеаз, которые действуют на пептидогликан (29, 30).Точная функция этих белков является актуальной темой исследований (31). Структурный анализ и анализ in vitro позволяют предположить, что по крайней мере некоторые из этих ферментов представляют собой d,d-карбоксипептидазы (расщепляют d-Ala с пятой позиции пептидного стебля) и d,d-эндопептидазы (расщепляют поперечные связи). Эти ферментативные активности, вероятно, будут нарушены/модифицированы в бактериях с синтетической клеточной стенкой. Однако из-за различий в профиле ферментов и общей архитектуре клеточной стенки ( Campylobacter является грамотрицательным) между микроорганизмами мы не можем в настоящее время делать выводы из сравнения.
Активность ферментов, разрушающих клеточную стенку, влияет на содержание синтетической клеточной стенки.
Ранее мы наблюдали образование ангидроNAM-сахарида на первичном сшитом d-AA муропептиде в E. coli (9). АнгидроНАМ представляет собой продукт реакции литических трансгликозилаз (ЛТ), ферментов, которые разрушают клеточную стенку путем негидролитического расщепления гликозидной связи между НАМ и НАГ с образованием продукта реакции ангидроНАМ (показан на рис. 2 D ). Активность LT доминирует у грамотрицательных бактерий и в образцах изолированных клеточных стенок 3.7% муропептидов содержат ангидро-NAM, который закрывает концы гликановых цепей после расщепления (32). Используя ЖХ-МС/МС, авторы картировали эту модификацию сахарида, присоединенного к пептидному стеблю, который не модифицируется d-AA. Мы не наблюдали образования ангидро-NAM на первичном несшитом муропептиде, содержащем d-AA, а также на пептидной основе сшитого муропептида, модифицированного d-AA. Это наблюдение предполагает, что LT не могут образовывать кепки ангидро-NAM на сахаридах NAM, примыкающих к нашим d-AA-модифицированным пептидным стеблям.Интересно, что в текущем исследовании мы наблюдали модификацию ангидро-NAM в том же самом положении (т. е. сахарид, прикрепленный к стеблю пептида, который не модифицировался d-AA сшитого муропептида) у B. subtilis (фиг. 2 Д ). Грамположительные бактерии преимущественно используют гидролитические мурамидазы для расщепления гликозидной связи между NAM и NAG (33), а соответствующие продукты реакции гидролиза служат «шапками» на концах их гликановых цепей (34).Активность LT у грамположительных бактерий менее выражена, и в изолированных образцах нативной клеточной стенки только 0,4% муропептидов содержат ангидро-NAM (18).
В E . coli мы предположили, что нерасщепляемая природа синтетической поперечной связи мешает экзолитическому процессингу (отщепление дисахарида от гликанового конца) LT, так что гликозидная связь, расположенная на C4 GlcNAc, не может быть расщеплена (рис. 5 ). А , I-III). Поскольку LT не способны разрушать клеточную стенку синтетических сшитых муропептидов, на синтетические сшивки наносится отпечаток ангидро-NAM, и содержимое синтетической клеточной стенки начинает накапливаться по мере того, как скорость образования синтетической клеточной стенки превосходит скорость деградации. (Инжир.5 А , IV ). Чтобы оценить, приводит ли нарушение активности LT синтетической клеточной стенкой к накоплению синтетических муропептидов, мы обработали E. coli 1 мМ 1a в течение 8 часов. После каждого 2-часового интервала мы собирали 50 мл культуры и проводили ЖХ-МС выделенного муропептида для определения содержания синтетической клеточной стенки (рис. 5 B и C ). Содержание синтетических клеточных стенок значительно увеличивалось от 0 до 4 часов после обработки; через 6 ч накопление замедляется, что может быть связано с гомеостазом, достигаемым на поздней стационарной фазе (рис.5 C и D ). Чтобы экспериментально подтвердить, что LT-активность E . coli нарушается как несшитыми, так и сшитыми муропептидами, содержащими d-AA, мы обработали клеток E. coli 1 мМ неканонического контрольного соединения O -метил-d-тирозина (структура в SI Приложение , рис. S14 A ), который структурно напоминает 1a , но является нереакционноспособным. Мы снова наблюдали картину накопления, которая напоминает таковую для культуры, обработанной 1a ( SI Приложение , рис.S14 B – G ), демонстрируя, что активность LT нарушается d-AA-модифицированными пептидными стеблями, даже если синтетическая поперечная связь не образуется.
Рис. 5.( A ) Упрощенное изображение предполагаемого пути деградации гликанов синтетически модифицированной клеточной стенки грамотрицательных бактерий E. coli с помощью расщепляющих гликан ферментов. Неканонические d-AA показаны желтыми кружками. Красные линии указывают на потенциальные участки разреза ферментов на каждой соответствующей панели.( B ) Кривая роста бактерий E. coli , обработанных соединением 1a (1 мМ). Серые кружки обозначают точки сбора роста, в которых рост был остановлен осаждением и замораживанием бактериальной культуры. ( C ) ВЭЖХ-кривая (абс. 205 нм) бактериальной клеточной стенки E. coli в каждой точке сбора. ( D ) Количественное определение муропептидов SnC и SC относительно общей концентрации муропептидов nC и SnC, а также C и SC соответственно.( E ) Упрощенное изображение предполагаемого пути деградации гликанов синтетически модифицированной клеточной стенки у грамположительных бактерий B. subtilis ферментами, расщепляющими гликан. ТП не показаны. ( F ) Кривая роста бактерий B. subtilis s, обработанных соединением 1a (1 мМ) ( G ) Кривая ВЭЖХ (абс. 205 нм) бактериальной клеточной стенки B. subtilis . ( H ) Количественное определение SnC и SC муропептидов B. subtilis .Обозначения для структур со стенками ячеек представлены на рис. 1.
In B . subtilis LT, вероятно, продуцируют продукты реакции, сходные с теми, что образуются в E. coli (рис. 5 E , I , II ). Однако гидролитические мурамидазы B . subtilis расщепляют и вырезают как нативную, так и синтетическую клеточную стенку, что должно приводить к меньшему накоплению синтетической клеточной стенки, чем в E. coli , где деградация более нарушена (рис.5 E , III , IV ). ВЭЖХ-анализ образцов клеточной стенки, выделенных из B. subtilis , в зависимости от времени показывает содержание клеточной стенки, аналогичное E. coli , через 2 часа после обработки. Однако у B. subtilis значительного накопления синтетической клеточной стенки в течение 2 ч не наблюдалось (рис. 5 G и H ). Следовательно, быстро достигается гомеостаз между включением и последующим образованием синтетических поперечных связей неканонических d-AA и удалением синтетической клеточной стенки гидролитическими мурамидазами.
Заключение.
Здесь мы показали, что клеточная стенка грамположительной палочковидной бактерии может быть модифицирована электрофильными неканоническими d-AA, которые образуют неферментативно катализируемые синтетические поперечные связи клеточной стенки в размере 19% от общего количества клеточных стенок. поперечные связи стенок живой бактерии. В В . subtilis , высокий уровень синтетических поперечных связей клеточной стенки, по-видимому, препятствует синтезу перегородки клеточной стенки, но не синтезу боковой стенки клеточной стенки, придавая бактериям удлиненную морфологию.Мы также начали выяснять ферментативную активность, нарушаемую синтетическими поперечными связями клеточной стенки, и результирующие эффекты на синтез и деградацию клеточной стенки. Способность изменять расположение синтетических поперечных связей в живых бактериях путем удаления генов ферментов биосинтеза клеточной стенки позволяет изучать очень разнообразные структуры клеточной стенки. Мы показали, что ненативные синтетические 4,3-поперечные связи и 5,3-поперечные связи, последняя из которых полностью отсутствует в природе, размещаются в клеточной стенке живых бактерий в больших количествах.Кроме того, недавно было показано, что синтетические 5,5-сшивки могут образовываться в клеточных стенках живых бактерий путем азид-алкинового циклоприсоединения, хотя для этой реакции требуется экзогенная медь и соответствующий медный лиганд (35). Вполне вероятно, что более обширные модификации могут быть сделаны в архитектуре клеточной стенки с использованием подобных подходов.
Автоклавные ячеистые бетонные блоки Продукт для каменной кладки AAC Alc Стеновой блок — Китай Более прочная структура и малая плотность AAC Блок, экологически безопасный AAC Блок продукт, который разработан в качестве строительного материала для наружных стен.Мы разработали внутренние стеновые блоки различной толщины, в том числе 150 мм, 200 мм, 240 мм, 250 мм, 300 мм, различная толщина может соответствовать различным требованиям архитектурного дизайна.
Ecotrend Блок для наружных стен из газобетона представляет собой разновидность легкого стенового материала. По сравнению с традиционными стеновыми материалами, он обладает превосходными сейсмическими характеристиками, высокой прочностью, не подвержен старению, высокой долговечностью, а также имеет хорошие характеристики теплоизоляции, звукоизоляции, огнеупорности, влагостойкости.
Это отличный материал для наружных стен зданий, широко используемый в различных наружных стенах зданий, особенно наружных стенах сборных зданий. Кроме того, он также широко используется в качестве стенового материала.
Автоклавный газобетонный блок (сокращенно AAC) представляет собой разновидность легкого сборного бетонного блока, он изготовлен из цемента, извести, летучей золы (или кварцевого песка) в качестве основного сырья и алюминиевого порошка в качестве воздухововлекающего агент. Это новый тип строительного материала с превосходными характеристиками, который производится в условиях высокой температуры, высокого давления и отверждения паром.
Отличительные преимущества:
(1) Малый вес:
Сухая плотность газобетонных блоков обычно составляет 400–700 кг/м³, что эквивалентно одной трети глиняного кирпича и одной четверти бетона, при этом также ниже, чем обычный бетон с легким заполнителем, если используются блоки AAC, можно сэкономить транспортные расходы, затраты на энергию и рабочую силу.
(2) Высокая прочность:
Газобетонные блоки Ecotrend имеют высокую точность размеров. Если для кладки используется тонкослойный раствор, коэффициент использования прочности значительно повышается.Прочность кладки составляет около 80 % прочности самого блока, что на 30 % прочнее глиняного кирпича. Кроме того, стальные стержни могут быть сконфигурированы в соответствии с проектными требованиями для удовлетворения различных потребностей.
(3) Сейсмостойкость:
Благодаря небольшому весу газобетонных блоков собственный вес зданий, в которых используются газобетонные блоки, значительно снижается, что значительно повышает сейсмостойкость здания.
(4) Долговечность:
Газобетонные блоки Ecotrend представляют собой разновидность силикатного материала, не подвержены старению и не поддаются атмосферным воздействиям.Это прочный сборный строительный материал, и его нормальный срок службы можно сравнить со всеми видами капитальных построек.
Прочность на компрессию (MPA / м³) | Средняя плотность (кг / м³) | |||
Среднее значение | Минимальное значение | |||
600*150*200 | ≥5.0 | ≥4.0 0≥4.0 | ≤625 (725) | |
600 * 200 * 200 | ≥5,0 | ≤625 (725) | ||
600 * 240 * 200 600 * 240 * 200 | ≥5.0 0 | ≥4.0 0 | ≤625 (725) | |
600 * 250 * 200 | ≥5,0 | ≥4,0 | ≤625(725) | |
600*300*200 | ≥5.0 | ≥4,0 | ≤625(725) | |
B06 | |||||||
Сухая плотность (кг / м³) | ≤425 | ≤525 | ≤625 | ||||
Прочность на сжатие (МПа/м³) | A2.0 | A5.0 | |||||
0,09 0 | 0.132 | ||||||
Сушильная усадка (мм / м) | 0.3 | 0.32 | |||||
Устойчивость к замораживанию | ≤3 | ≤3 | 2.1 | ||||
Прочность после заморозки (MPA) | ≥2,0 | ||||||
Умягчающий коэффициент | 0.88 | ||||||
Проникновение Сопротивление | 88.3 | 88.3 | |||||
Огнестойкость (час) | 40005 | 4hour (толщина блока стенки: 100 мм) | |||||
Средняя звукоизоляция (БД) | 40.8 (толщина блока: 100 мм, с двумя сторонами) | ||||||
Длина ± 2, ширина ± 1, толщина ± 1 | |||||||
Процесс продукта
Автоклавный газобетон (сокращенно AAC) — это разновидность легкого сборного железобетона, он был изобретен в середине 1920-х годов и развивался около 100 лет. С 1980 года производство газобетонных блоков продолжает расти во всем мире, и они все чаще используются разработчиками, архитекторами, строителями по всему миру, особенно для сборных домов, что соответствует тенденции развития строительной отрасли.
Технические характеристики и стандарт качества блока ECOTREND соответствуют требованиям национального стандарта GB11968-2006 на блоки из ячеистого бетона автоклавного твердения. Подходит для гражданских и промышленных зданий в районах с сейсмичностью менее 7 градусов, возводимых из ненесущих стеновых материалов по технологии сухой тонкослойной кладки; он подходит для сохранения тепла и энергосбережения зданий, которые широко используются в жилых домах, офисных зданиях, больницах, гостиницах, школах и других гражданских, общественных и промышленных зданиях.
Применение продукта
Упаковка и транспортировка
Ecotrend представила передовую в мире линию по производству газобетона для производства газобетонных блоков и панелей ALC (панели ALC относятся к сборным строительным материалам). Годовая производственная мощность компании составляет 400 000 кубических метров, что является крупнейшим и единственным предприятием по производству панелей ALC с экологичными сборными строительными материалами в Камбодже. Так как он находится всего в 20 минутах езды от порта Сианук Вилль, Ecotrend обладает уникальным географическим преимуществом для экспорта.
Профиль компании
Ecotrend (Камбоджа) New Building Materials Development Co., Ltd. – это предприятие с китайскими инвестициями, основанное в Королевстве Камбоджа, специализирующееся на разработке, производстве и экспорте экологически чистых, энергоэффективных новые строительные материалы — продукты автоклавного газобетона (автоклавного легкого бетона) (известные как продукты AAC или ALC). Компания Ecotrend расположена в Особой экономической зоне Сианук-Вилль, Камбоджа. Компания занимает земельный участок площадью около 25000 квадратных метров; и общий объем инвестиций более 14 миллионов долларов.У нас есть передовые технологии и сертификаты соответствия стандартам
Почему выбирают нас?
1). Наша годовая производственная мощность составляет 400 000 кубических метров , что является ведущим производителем в Юго-Восточной Азии.
2). Удобный транспорт: наша фабрика находится по адресу недалеко от порта Сианук-Вилль , что всего в 20 минутах езды от порта .
3). Передовые технологии производства: мы представили полностью автоматические производственные линии в мире для производства блоков AAC и панелей ALC (панели ALC относятся к сборным строительным материалам).
4). Наша миссия: поставлять газобетонные изделия для ваших проектов, которые будут соответствовать вашим требованиям с точки зрения качества, количества, спецификаций, технических характеристик и
по конкурентоспособной цене.
FAQ
Q: Какая связь между Hangzhou Huafei Express Import and Export Co., Ltd и Ecotrend (Камбоджа) New Building Materials Development Co., Ltd?
Ответ: У них один и тот же акционер. HuaFei Express — это маркетинговый центр, расположенный в Китае.Ecotrend является производителем продукции AAC/ALC в Камбодже.
В: С кем мы подпишем контракт?
A: Обычно с Ecotrend для продуктов AAC/ALC, и отгрузка будет осуществляться непосредственно из Камбоджи. Если вы хотите купить аксессуары для продуктов ALC, вы подписываете контракт с Huafei Express.
В: В какой порт вы будете экспортировать свою продукцию?
A: Для продуктов AAC/ALC мы будем экспортировать из порта Сианук-Вилле, Камбоджа; для аксессуаров или других сопутствующих товаров, мы будем экспортировать из Шанхая или порта Нинбо, Китай.
В: какое у вас время выполнения заказа?
A: Обычно в течение 15-20 дней после получения депозита или аккредитива.
В: Каковы ваши условия оплаты?
A: 50% предоплата банковским переводом и 50% баланса банковским переводом перед погрузкой; Аккредитив также принимается.
Безумное движение строительных блоков – нуклеотидные сахара находят неожиданные пути в клеточные стенки | Журнал экспериментальной ботаники
Ряд недавних исследований, в том числе новая статья Чжао и др. (2018) пролил новый свет на важность клеточной топологии в контроле потока нуклеотидных сахаров к месту назначения. Это захватывающее время, так как быстро прогрессирующее выяснение запутанного потока углеродных тел посредством метаболизма нуклеотидных сахаров и межорганеллярного транспорта означает, что мы приближаемся к пониманию принципов управления биосинтезом полисахаридов растительной клеточной стенки.
Наука о растениях приближается к видению клеточных стенок, построенных «соответствующих назначению» (Johnson et al., 2017). Сборка углеводов из моносахаридов включает стереоспецифический перенос специфических (ди)фосфонуклеотид-активированных сахаров на специфические акцепторные молекулы с помощью гликозилтрансфераз, которые, по оценкам, кодируются более чем 500 локусами Arabidopsis (Hansen et al. , 2012). По крайней мере, 16 нуклеотидных сахаров образуются в сети взаимопревращения нуклеотидных сахаров, и многие из задействованных ферментов в настоящее время клонированы и охарактеризованы (Reiter and Vanzin, 2001; Seifert, 2004; Reiter, 2008; Bar-Peled and O’Neill, 2011). .Совсем недавно были идентифицированы локализованные по Гольджи переносчики нуклеотидных сахаров (Temple et al. , 2016). Но как метаболизм и транспорт нуклеотидных сахаров связаны с биосинтезом углеводов в живых растениях?
Регулирование флюса подложки
Большинство нуклеотидных сахаров производится из UDP-D-глюкозы (UDP-Glc) в цитозоле (вставка 1). Этот центральный предшественник углеводов превращается в УДФ-L-рамнозу, УДФ-галактозу и УДФ-D-глюкуроновую кислоту (УДФ-GlcA), причем последняя превращается в УДФ-D-апиозу и УДФ-D-ксилозу (УДФ-ксил). .Только фермент, который генерирует UDP-D-галактуроновую кислоту (GalA) из UDP-GlcA, локализован исключительно по методу Гольджи. Напротив, два фермента, взаимопревращающие UDP-сахар, существуют в двух формах, локализованных в топологически разделенных компартментах — одна форма в цитозоле, а другая в аппарате Гольджи. Причина этой специфичной для растений двойственности не сразу очевидна.
Вставка 1. Метаболические пути УДФ-ксила и УДФ-Ара f
Коричневые стрелки указывают на судьбу в planta углеродных тел, превращающихся в ксилозу, ассоциированную с клеточной стенкой: глюкуроноксилан (GX), ксилоглюкан ( XG), ксилогалактуронана (XGA) и сложного N-связанного гликана (CGL).Зеленые стрелки указывают на судьбу углеродных тел, заканчивающихся арабинофуранозой, связанной с полимерами клеточной стенки: рамногалактуронан I (RG I) и RG II, белок арабиногалактана (AGP) и экстенсин (EXT). Красная стрелка указывает на поток от UDP-Glc к UDP-GlcA, который является общим для арабинозных и ксилозных телец и опосредуется дегидрогеназой UDP-Glc (UGD). Тонкие серые стрелки указывают на потенциальные метаболические пути, которые, по-видимому, не используются в planta . Участвующие ферменты: UDP-Glc-дегидрогеназа (UGD), UDP-ксилсинтаза (UXS), UDP-ксилозо-4-эпимераза (UXE), биспецифическая UDP-Glc/UDP-Xyl-4-эпимераза (UGE), арабинокиназа (AK), UDP -сахарная пирофосфорилаза (USP) и обратимо гликозилированный полипептид (RGP).Транспортеры нуклеотидных сахаров представляют собой переносчик UDP-Xyl (UXT), переносчик UDP-уроновой кислоты (UUAT), гипотетический облегчающий экспорт UDP-Ara p (серый) и переносчик UDP-арабинофуранозы (UAfT). Символы для «ксилозилтрансферазы» (XT) и «арабинозилтрансферазы» (AT) в общем представляют разные гликозилтрансферазы. Обратите внимание, что пути, ведущие к UDP-галактозе, UDP-L-рамнозе и UDP-галактуроновой кислоте или любым GDP-сахарам, не показаны.
Первым примером является биосинтез UDP-Xyl, предшественника ксиланов (York and O’Neill, 2008), ксилоглюкана, рамногалактуронана II (RG II) и ксилогалактуронана (Caffall and Mohnen, 2009).UDP-Xyl synthase (UXS) катализирует декарбоксилирование UDP-GlcA в UDP-Xyl с генами UXS3 , -5 и -6 , кодирующими цитозольный UXS и UXS1 , -2 3 4 локусов, кодирующих локализованные по Гольджи изоформы. Поскольку UDP-Xyl требуется исключительно внутри Golgi, три цитозольные изоформы UXS кажутся избыточными. Что еще более важно, чтобы предоставить цитозольному пулу UDP-Xyl доступ к месту биосинтеза углеводов, необходим Golgi-localized транспортер UDP-Xyl.Действительно, три транспортера UDP-Xyl (UXT1-3) были изобретательно идентифицированы с помощью биохимического анализа, а мутант uxt1 показал дефект в структуре и распространенности глюкуроноксилана (GX). Этот фенотип предполагает, что биосинтез полисахаридов каким-то образом зависит от цитозольного UDP-Xyl, несмотря на присутствие функционального UXS в Golgi. Однако наблюдалось лишь относительно незначительное снижение общего содержания ксилозы в клеточной стенке у uxt1 , и у мутанта не затрагивались никакие другие углеводы, кроме GX.Более того, двойной мутант uxt2 uxt3 был фенотипически нормальным (Ebert et al. , 2015).
Этот ключевой документ Ebert et al. (2015) открыл важные вопросы для понимания того, как регулируется поток субстрата в planta . Отражает ли GX-специфический дефект в uxt1 канализацию субстрата от UXT1 к GX-специфическим ксилозилтрансферазам? И наоборот, , является относительно тонким общим фенотипом uxt1 из-за генетической избыточности в транспорте UDP-ксилозы или действия локализованного по Гольджи UXS? Чжао и др. (2018) отвечают на эти вопросы, показывая, что у тройного мутанта uxt1 uxt2 uxt3 сильно нарушены рост растений и утолщение вторичной клеточной стенки, а помимо GAX дефектен и ксилоглюкан. Хотя это и не противоречит гипотезе канализации субстрата, их статья убедительно демонстрирует решающее значение цитозоля для транспорта по Гольджи UDP-Xyl для множества ксилозосодержащих углеводов. Это еще раз подтверждает предыдущие исследования с тройными мутантами, сравнивающие две формы UXS, которые показали, что цитозольный UXS, а не локализованный в Гольджи фермент, влияет на включение ксилозы в полисахариды клеточной стенки (Kuang et al., 2016; Чжун и др. , 2017).
Механическая основа разделения
Какова может быть механистическая основа для этого кажущегося разделения потоков метаболитов (вставка 1)? Общей чертой как UXS, так и UDP-Glc-дегидрогеназы (UGD), которая генерирует UDP-GlcA, является то, что оба фермента ингибируются UDP-Xyl (Harper and Bar-Peled, 2002; Pattathil et al. , 2005; Klinghammer). и Тенхакен, 2007). Следовательно, онтогенетически активная регуляция цитозольного UXS в начале образования вторичной клеточной стенки будет увеличивать цитозольный UDP-Xyl, тем самым ингибируя UGD по обратной связи.Ингибирование UGD будет подавлять биосинтез как UDP-GalA, так и, через локализованный в Гольджи биосинтез UDP-Xyl, UDP-L-арабинозы (UDP-Ara), которые в основном необходимы во время биосинтеза первичной клеточной стенки. Но если ксилозосодержащие углеводы получают UDP-Xyl из цитозольного UXS, какова биологическая функция формы Гольджи? В двух генетических исследованиях не удалось установить четкую роль UXS1, -2 и -4 (Kuang et al. , 2016; Zhong et al. , 2017).Однако, поскольку локусы в основном экспрессируются в тканях, которые претерпевают первичное формирование клеточной стенки (Kuang et al. , 2016), было бы интересно дополнительно исследовать углеводно-химический фенотип uxs1 uxs2 uxs4 в активно расширяющихся тканях.
Косвенные доказательства важности локализованного по Гольджи UXS в обеспечении субстрата для UXE и, в конечном счете, для арабинозилтрансфераз были получены в результате обнаружения того, что транспортер для UDP-GlcA, называемый UUAT1, важен для содержания арабинозы в клеточных стенках (Saez-Aguayo). и др., 2017). Оказавшись внутри аппарата Гольджи, UDP-GlcA либо превращается в UDP-GalA с помощью UDP-глюкуроновой кислоты 4-эпимеразы (GAE), либо в UDP-Xyl с помощью UXS. Из UDP-Xyl UDP-Xyl 4-эпимераза (UXE) генерирует UDP-Ara, и, что особенно важно, мутант uxe1 показал резкое снижение содержания арабинозы в клеточной стенке (Burget and Reiter, 1999; Burget et al. , 2003).
Пентагональная арабинофураноза, гексагональная пираноза
Другим важным аспектом биосинтеза UDP-Ara является то, что пентагональная арабинофураноза (Ara f ), а не гексагональная пираноза (Ara p ) форма L-арабинозы обнаружена в большинстве полимеров клеточных стенок.Однако для эффективного превращения продукта UXE UDP-Ara p в UDP-Ara f требуется мутаза. Эта мутаза представляет собой белковый комплекс, кодируемый локусами REVERSIBLY GLYCOSYLATED POLYPEPTIDE1 , -2 и — 5 ( RGP1, -2 и -5 ) и частично связан с цитозольной поверхностью G. Нокдаун RGP1 и RGP2 привел к массивным дефектам роста и почти полной потере арабинозы клеточной стенки (Rautengarten et al., 2011). Недавно идентифицированный транспортер UDP-арабинофуранозы UAfT (Rautengarten et al. , 2017) направляет UDP-Ara f обратно в аппарат Гольджи, где он является субстратом для множества арабинозилтрансфераз. Однако потенциальные пути метаболита в связанный с клеточной стенкой Ara f на этом не заканчиваются. Ожидается, что
UDP-Ara p , экспортируемый из аппарата Гольджи в цитозоль, будет смешиваться с UDP-Ara p , образующимся из свободной L-арабинозы в процессе расщепления углеводов с помощью арабинокиназы и УДФ-сахарной пирофосфорилазы (Geserick and Tenhaken, 2013; Behmüller). и др., 2016). Этот спасительный путь является механистическим объяснением реверсии фенотипа uxe1 с помощью экспериментально примененного L-Ara (Burget and Reiter, 1999). И еще один путь потенциально ведет к UDP-Ara f. Цитозольная бифункциональная UXE/UDP-глюкозо-4-эпимераза (изоформы UGE1 и UGE3) (Kotake et al. , 2009) фактически должна генерировать цитозольный UDP-Ara p из обильного поступления UDP-Xyl цитозольным УСС. Однако этому потенциальному варианту противоречит выраженный дефицит арабинозы в клеточных стенках мутанта uxe1 (Burget and Reiter, 1999) и отсутствие фенотипа у двойного мутанта uge1 uge3 (Rösti et al., 2007). Одним из возможных объяснений может быть то, что точная коэкспрессия на уровне как мРНК, так и белка необходима для нормального потока метаболитов во время роста и развития. Другой возможностью объяснить наблюдаемую специфичность является субстратный канал между соседними ферментами и транспортерами, собранными в комплекс. В любом случае, хотя метаболические пути нуклеотидных сахаров кажутся капризными, они, несомненно, представляют собой гибкую и эффективную систему доставки, закаленную миллиардами лет эволюции.
Благодарности
Автор поддерживается грантом Австрийского научного фонда FWF номер I1182-B22.
Каталожные номера
Бар-Пеледь
М
,О’Нил
Массачусетс
.2011
.Образование растительных нуклеотидных сахаров, взаимная конверсия и утилизация путем переработки сахара
.Ежегодный обзор биологии растений
62
,127
–155
.Бемюллер
Р
,Кавкова
E
,Düh
S
,Huber
CG
,Tenhaken
R
.2016
.Роль арабинокиназы в токсичности арабинозы у растений
.Журнал завода
87
,376
–390
.Бергет
ЭГ
,Райтер
WD
.1999
.Мутант арабидопсиса mur4 частично дефектен в синтезе de novo уридиндифосфо L-арабинозы
.Физиология растений
121
,383
–389
.Бергет
ЭГ
,Verma
R
,Mølhøj
M
,Reiter
WD
.2003
.Биосинтез L-арабинозы в растениях: молекулярное клонирование и характеристика Гольджи-локализованной UDP-D-ксилозо-4-эпимеразы, кодируемой геном MUR4 арабидопсиса
.Растительная клетка
15
,523
–531
.Каффолл
КХ
,Монен
D
.2009
.Структура, функция и биосинтез пектиновых полисахаридов клеточной стенки растений
.Исследование углеводов
344
,1879
–1900
.Эберт
Б
,Rautengarten
C
,Guo
X
и др.2015
.Идентификация и характеристика локализованного по Гольджи семейства переносчиков UDP-ксилозы из Arabidopsis
.Растительная клетка
27
,1218
–1227
.Гезерик
С
,Тенхакен
Р
.2013
.UDP-сахарная пирофосфорилаза необходима для рециркуляции арабинозы и ксилозы, а также требуется во время вегетативного и репродуктивного роста арабидопсиса
.Журнал завода
74
,239
–247
.Хансен
СФ
,Harholt
J
,Oikawa
A
,Scheller
HV
.2012
.Растительные гликозилтрансферазы за пределами CAZy: взгляд на семейства DUF
.Frontiers in Plant Science
3
,59
.Харпер
г. н.э.
г. ,Бар-Пелед
М
.2002
.Биосинтез УДФ-ксилозы. Клонирование и характеристика нового семейства генов Arabidopsis, UXS, кодирующих растворимые и предполагаемые мембраносвязанные изоформы декарбоксилазы UDP-глюкуроновой кислоты
.Физиология растений
130
,2188
–2198
.Джонсон
КЛ
,Gidley
MJ
,Bacic
A
,Doblin
MS
.2017
.Биомеханика клеточных стенок: решаемая задача манипулирования клеточными стенками растений, «соответствующими своему назначению»
!Актуальное мнение в области биотехнологии
49
,163
–171
.Клингхаммер
М
,Тенхакен
Р
.2007
.Полногеномный анализ семейства генов UDP-глюкозодегидрогеназы у арабидопсиса, ключевого фермента полисахаридов матрикса в клеточных стенках
.Журнал экспериментальной ботаники
58
,3609
–3621
.Котаке
Т
,Takata
R
,Verma
R
и др.2009
.Бифункциональные цитозольные УДФ-глюкозо-4-эпимеразы катализируют взаимопревращение между УДФ-D-ксилозой и УДФ-L-арабинозой в растениях
.Биохимический журнал
424
,169
–177
.Куанг
Б
,Чжао
X
,Чжоу
C
и др.2016
.Роль декарбоксилазы УДФ-глюкуроновой кислоты в биосинтезе ксилана у арабидопсиса
.Молекулярная установка
9
,1119
–1131
.Паттатил
С
,Харпер
АД
,Бар-Пелед
М
.2005
.Биосинтез УДФ-ксилозы: характеристика мембраносвязанного AtUxs2
.Планта
221
,538
–548
.Раутенгартен
С
,Birdseye
D
,Pattathil
S
и др.2017
.Сложный путь доставки УДФ-арабинозы в Гольджи растений
.Proceedings of the National Academy of Sciences, USA
114
,4261
–4266
.Раутенгартен
С
,Ebert
B
,,
T
,T
,T
CJ,CJ
,ISHII
T
,MUKHOPADHYAY
A
,USADEL
B
,Scheller
HV
.2011
.Взаимное превращение УДФ-арабинопиранозы и УДФ-арабинофуранозы необходимо для развития растений у арабидопсиса
.Растительная клетка
23
,1373
–1390
.Райтер
ВД
.2008
.Биохимическая генетика реакций взаимопревращения нуклеотидов и сахаров
.Текущее мнение по биологии растений
11
,236
–243
.Райтер
ВД
,Ванзин
ГФ
.2001
.Молекулярная генетика путей взаимопревращения нуклеотидов и сахаров в растениях
.Молекулярная биология растений
47
,95
–113
.Рости
Дж
,Barton
CJ
,ALBRECHT
S
,S
,,
P
,Pauly
M
,Findlay
K
,Roberts
K
,Seifert
GJ
.2007
.Изоформы UDP-глюкозо-4-эпимеразы UGE2 и UGE4 совместно обеспечивают UDP-галактозу для биосинтеза клеточной стенки и роста Arabidopsis thaliana
.Растительная клетка
19
,1565
–1579
.Саес-Агуайо
С
,Rautengarten
C
,Temple
H
и др.2017
.UUAT1 представляет собой переносчик УДФ-уроновой кислоты, локализованный в точке Гольджи, который модулирует полисахаридный состав слизи семян арабидопсиса
.Растительная клетка
29
,129
–143
.Зайферт
ГДж
.2004
.Взаимопревращения нуклеотидных сахаров и биосинтез клеточной стенки: как вывести внутреннее наружу
.Текущее мнение по биологии растений
7
,277
–284
.Храм
Х
,Саес-Агуайо
С
,Рейес
ФК
,Орельяна
А
.2016
.Внутри и снаружи: топологические проблемы в биосинтезе клеточной стенки растений и роль переносчиков нуклеотидных сахаров
.Гликобиология
26
,913
–925
.Йорк
ВС
,О’Нил
Массачусетс
.2008
.Биохимический контроль биосинтеза ксилана — какой конец выше
?Текущее мнение по биологии растений
11
,258
–265
.Чжао
х
,Лю
N
,Шан
N
и др.2018
.Три переносчика UDP-ксилозы (UXT) участвуют в биосинтезе ксилана путем переноса цитозольной UDP-ксилозы в полость Гольджи у Arabidopsis
.Журнал экспериментальной ботаники
69
,1125
–1134
.Чжун
Р
,Teng
Q
,Haghighighat
M
,юаней
Y
,Furey
,
,
ST
,Dasher
RL
,YE
ZH
.2017
.Локализованные в цитозоле синтазы УДФ-ксилозы обеспечивают основной источник УДФ-ксилозы для биосинтеза ксилана и ксилоглюкана
.Физиология растений и клеток
58
,156
–174
.© Автор(ы), 2018 г. Опубликовано Oxford University Press от имени Общества экспериментальной биологии.
Это статья в открытом доступе, распространяемая на условиях лицензии Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), что разрешает неограниченное повторное использование, распространение и воспроизведение на любом носителе при условии правильного цитирования оригинальной работы.Как антибиотики убивают бактериальные клетки, но не клетки человека?
Гарри Мобли, заведующий кафедрой микробиологии и иммунологии Медицинской школы Мичиганского университета, дает такой ответ.
Чтобы быть полезными при лечении инфекций человека, антибиотики должны избирательно нацеливаться на бактерии для уничтожения, а не на клетки человека-хозяина.Действительно, современные антибиотики действуют либо на процессы, которые являются уникальными для бактерий, такие как синтез клеточных стенок или фолиевой кислоты, либо на специфические для бактерий мишени в процессах, которые являются общими как для клеток бактерий, так и для клеток человека, включая репликацию белков или ДНК. . Ниже приведены некоторые примеры.
Большинство бактерий образуют клеточную стенку, частично состоящую из макромолекулы, называемой пептидогликаном, которая сама состоит из аминосахаров и коротких пептидов. Клетки человека не производят и не нуждаются в пептидогликанах.Пенициллин, один из первых широко используемых антибиотиков, предотвращает последнюю стадию сшивания или транспептидации при сборке этой макромолекулы. В результате очень хрупкая клеточная стенка разрывается, убивая бактерию. Человеку-хозяину не причиняется никакого вреда, потому что пенициллин не подавляет никаких биохимических процессов, происходящих внутри нас.
Бактерии также могут быть избирательно уничтожены путем воздействия на их метаболические пути. Сульфонамиды, такие как сульфаметоксазол, аналогичны по структуре парааминобензойной кислоте, соединению, имеющему решающее значение для синтеза фолиевой кислоты.Всем клеткам требуется фолиевая кислота, и она может легко диффундировать в клетки человека. Но витамин не может проникнуть в бактериальные клетки, поэтому бактерии должны вырабатывать свои собственные. Серосодержащие препараты, такие как сульфаниламиды, ингибируют в этом процессе важнейший фермент — дигидроптероатсинтазу. Как только процесс остановлен, бактерии больше не могут расти.
Другой вид антибиотиков — тетрациклин — также подавляет рост бактерий, останавливая синтез белка. И бактерии, и люди осуществляют синтез белка на структурах, называемых рибосомами.Тетрациклин может проникать через мембраны бактерий и накапливаться в высоких концентрациях в цитоплазме. Затем тетрациклин связывается с единственным участком рибосомы — 30S (меньшей) субъединицей рибосомы — и блокирует ключевое взаимодействие РНК, что отключает удлиняющуюся белковую цепь. Однако в клетках человека тетрациклин не накапливается в достаточных концентрациях, чтобы остановить синтез белка.
Точно так же репликация ДНК должна происходить как в бактериях, так и в клетках человека. В каждом из них процесс достаточно различен, поэтому антибиотики, такие как ципрофлоксацин — фторхинолон, известный своей активностью против бациллы сибирской язвы, — могут целенаправленно воздействовать на фермент, называемый ДНК-гиразой в бактериях.Этот фермент расслабляет плотно намотанную хромосомную ДНК, тем самым обеспечивая возможность репликации ДНК.